|
||||
|
|
Глава 2. На дальних подступах: приматы, обезьяны, гоминоиды Происхождение приматов Появление на эволюционной арене первых приматов приходится на рубеж мезозойской и кайнозойской эр, и это не случайно. Дело в том, что в конце мелового периода, завершающего мезозой, с лица земли исчезли господствовавшие дотоле на суше и в воде гигантские рептилии (динозавры, ихтиозавры, плезиозавры и т. д.), и в результате освободилось множество местообитаний и экологических ниш. Млекопитающие, скромно прозябавшие в течение десятков миллионов лет в тени пресмыкающихся, наконец, вышли на «оперативный простор» и начали активно заполнять образовавшиеся пустоты. Освоение разных экологических ниш вело к тому, что в поведении, физиологии и анатомии даже близкородственных групп накапливалось всё больше специфических черт, и их эволюционные пути в итоге расходились всё дальше. Следствием этого процесса, именуемого на языке биологов адаптивной радиацией, стало формирование в конце мелового периода и в начале палеогена множества новых видов, родов, семейств и отрядов животных. Очень интересен вопрос о том, как сложилась бы дальнейшая история жизни на Земле, если бы не массовое вымирание биоты на рубеже двух последних геологических эр. Этот вопрос не столь уж бессмыслен, как может показаться на первый взгляд, поскольку не исключено, что вымирание объясняется во многом случайными причинами, а среди тех, кого оно затронуло, могли быть и претенденты на продвижение в категорию мыслящих существ. Согласно широко известной и неплохо обоснованной гипотезе, исчезновение динозавров в конце мелового периода было вызвано катастрофой космического происхождения, т. е. падением крупного метеорита, повлёкшим за собой резкое изменение климата (его сравнивают иногда с эффектом «ядерной зимы»). Некоторые исследователи допускают, что не произойди этот катаклизм, нарушивший естественный, органичный ход развития земной природы, то нашей планетой сейчас владели бы ящеры, и разум появился бы не в своей нынешней материальной оболочке, а в мозгу каких-то из этих животных, например, целурозавров.[1] Конечно, это не более чем гипотеза, к тому же гипотеза непроверяемая, но в принципе в ней нет ничего невероятного, и она хорошо иллюстрирует идею потенциальной многовариантности эволюционного процесса. Вернёмся, однако, к нашим приматам. Согласно некоторым расчётам, основанным на количестве известных их видов (современных и ископаемых) и средней продолжительности «жизни» вида (1 миллион лет), первые представители отряда должны были существовать уже 80 млн лет назад, но большинству специалистов такая древность кажется маловероятной, поскольку значительно превышает возраст всех имеющихся ископаемых находок. Самые ранние из этих находок происходят из палеоценовых отложений и попадают в хронологический диапазон от 55 до 60 млн лет назад (см. рис. 2). Начальные стадии эволюции приматов изучены ещё недостаточно, и проблема происхождения отряда далека от окончательного разрешения. Ни его генеалогические корни, ни место появления пока точно не выявлены. Правда, практически не вызывает сомнений, что предками приматов были какие-то представители отряда насекомоядных (Insectivora), но, вместе с тем, среди имеющихся сейчас ископаемых находок нет таких, которые с полной уверенностью могли бы рассматриваться в качестве звена, связующего две эти группы животных. Обычно в качестве исходной предковой группы, промежуточной между насекомоядными, с одной стороны, и приматами, с другой, рассматривают род пургаториус (Purgatorius), известный по немногочисленным костям, происходящим из позднемеловых отложений Северной Америки. Ископаемые останки представителей этого рода и некоторых других, близких им, форм позволяют более или менее уверенно судить об облике и некоторых особенностях поведения древнейших приматов. Согласно существующим реконструкциям, это были мелкие (от приблизительно ста граммов до нескольких килограммов весом) насекомоядные и частично, по-видимому, растительноядные зверьки. Они вели преимущественно древесный образ жизни и, в отличие от своих предков, уже имели приспособленные к захвату конечности с относительно длинными фалангами пальцев и плоскими ногтями вместо когтей. За вычетом последней особенности внешне они, возможно, больше всего походили на современных тропических белок, и лишь благодаря специфическому строению зубов опознаются сейчас как приматы. Судя по географии находок, в палеоцене (65–54 млн лет назад) представители нового отряда обитали, главным образом, в Северной Америке и Западной Европе, соединённых в то время широкой полосой суши (рис. 3). Кроме того, отдельные ископаемые сопоставимой древности были обнаружены в Южной и Восточной Азии и в Африке, которые в начале кайнозойской эры также имели несколько иные очертания, нежели сейчас. 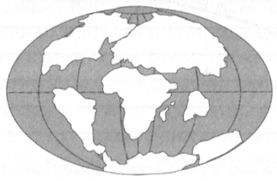
Первые обезьяны В раннем эоцене (54–45 млн лет назад) в пределах отряда приматов выделяется уже множество семейств, родов и видов, среди которых есть и предки современных лемуров и долгопятов. Обычно этих ранних полуобезьян делят на лемуриформных (лемуры и их предки) и тарзииформных (долгопяты и их предки). Не позднее 40 млн лет назад, т. е., скорее всего, в середине или даже начале эоцена, происходит, судя по находкам в Восточной Азии и Северной Африке, обособление линии высших приматов — антропоидов (Anthropoidea), или, иначе говоря, собственно обезьян (рис. 4). 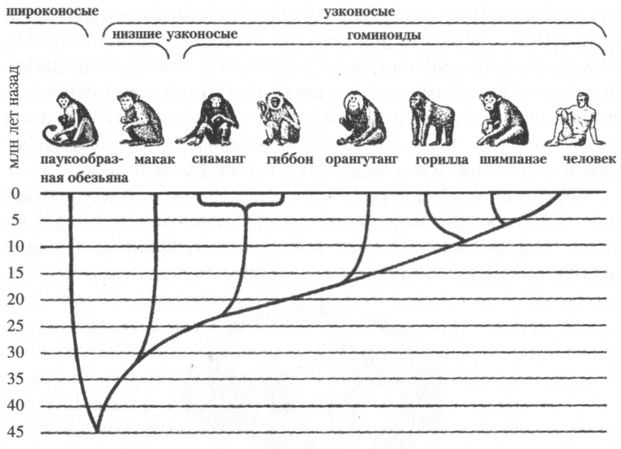
Следует иметь в виду, что в отечественной литературе термин антропоиды часто применялся, а иногда применяется и сейчас, для обозначения человекообразных обезьян. Однако такое его употребление, хотя и находит некоторое оправдание в этимологии самого слова «антропоиды» (оно происходит от греческого ???????? — человек и в буквальном переводе означает «человекоподобные»), нежелательно и ведёт к путанице. Антропоиды, согласно общепринятой зоологической номенклатуре, — это все обезьяны вообще, а для отдельного обозначения человекообразных обезьян существует и используется во всём мире термин «гоминоиды» (от латинского homo — человек). Вопрос о том, где появились первые обезьяны, пока не решён. На роль их прародины претендуют Африка и Восточная Азия, но сделать выбор между этими регионами при нынешнем состоянии наших знаний не представляется возможным. В последние годы некоторые важные находки, способные пролить свет на проблему происхождения антропоидов, были сделаны на территории Китая, Бирмы и сопредельных с ними стран, хотя и Африка, наверняка, не сказала ещё своё последнее слово в споре континентов, и здесь палеонтологов тоже ждут новые открытия. Уже первые обезьяны заметно отличались от низших приматов, или, иначе говоря, полуобезьян многими чертами строения зубной системы и глазничной области черепа, свидетельствующими об изменении характера рациона в сторону большей растительноядности (точнее, фругиворности, т. е. питания фруктами) и переходе от ночного образа жизни к дневному. С этими особенностями в их поведении связано и то обстоятельство, что зрение у них развито гораздо лучше, чем у большинства животных, а обоняние, наоборот, утратило остроту. По последним двум признакам к обезьянам близки долгопяты, на основании чего обе группы объединяют иногда в подотряд гаплорин (????? по-гречески означает «простой», а ?????[2] — «нос»). Более важно, однако, что антропоидам присуща такая черта, как относительно большой размер мозга, чем долгопяты похвастаться не могут. У долгопятов, как и у остальных приматов, объём мозга примерно такой же, как у других животных аналогичного размера, тогда как у обезьян он в среднем в два раза больше, чем следовало бы ожидать от млекопитающих их «весовой категории». Большинство палеонтологов выводят антропоидов из тарзииформных приматов, меньшинство — из лемуриформных, а, кроме того, высказывается предположение, что они могут представлять собой независимую ветвь, берущую начало непосредственно от общего для всех приматов исходного ствола. Впоследствии эта ветвь разделяется на две: широконосых обезьян (Platyrrhini), живущих сейчас только в Южной Америке, и узконосых (Catarrhini), населяющих Старый Свет. Наиболее древние кости широконосых обнаружены в Боливии, в отложениях возрастом около 25 млн лет, а вторых — в Северной Африке и на Аравийском полуострове, где они залегали в геологических слоях, сформировавшихся 31–35 млн лет назад, а частично, возможно, и ранее. Предки широконосых, бывшие, по-видимому, выходцами из Африки, могли случайно попасть в Южную Америку на естественных «плотах» из оказавшихся в воде растений. Сколь бы малой ни казалась вероятность благополучного исхода подобного путешествия, случаи такого рода, видимо, всё же происходили. Помимо предков широконосых обезьян, аналогичный путь проделали, как считается, и некоторые африканские грызуны. Почти всё, что мы знаем сегодня о ранних стадиях эволюции узконосых обезьян, стало известно благодаря находкам в Файюмской впадине, расположенной к западу от Нила на территории Египта. Это крупнейшее местонахождение среднеолигоценовой фауны дало останки множества видов приматов. Они происходят из отложений геологической формации Джебел Катрани, относящихся к периоду от 31 до 35 млн лет назад. Наибольший интерес среди обезьян, кости которых были найдены в Файюме, имеют для нас так называемые проплиопитециды, выделяемые обычно как надсемейство. К числу проплиопитецид относят роды проплиопитека (Propliopithecus) и египтопитека (Aegyptopithecus). В них многие исследователи видят наиболее вероятных предков современных узконосых обезьян, в том числе и человекообразных. Происхождение и эволюция человекообразных обезьян Примерно на рубеже олигоцена и миоцена (23 млн лет назад), или чуть раньше (см. рис. 2) происходит разделение дотоле единого ствола узконосых обезьян на две ветви: церкопитекоидов, или собакоподобных (Cercopithecoidea) и гоминоидов, т. е. человекообразных (Hominoidea). Это разделение, по всей видимости, во многом было обусловлено тем, что часть узконосых (предки церкопитекоидов) перешла к питанию листьями, тогда как другая часть (предки гоминоидов) сохранила верность фруктовой диете. Различия в меню сказались, в частности, на строении зубов, что чрезвычайно важно для палеонтологов, поскольку именно зубы составляют бо?льшую часть ископаемых находок. Поверхность жевательных зубов церкопитекоидов имеет характерный, только им присущий узор, образуемый четырьмя бугорками. На зубах же человекообразных обезьян пять округлых бугорков, разделённых У-образной бороздкой — так называемый «узор дриопитека» (рис. 5).  
Церкопитекоидов, представленных единственным, но очень многочисленным семейством мартышкообразных, часто называют низшими узконосыми обезьянами, а гоминоидов — высшими. Помимо особенностей формы зубов, гоминоидов отличает от низших узконосых обезьян также отсутствие хвоста, более короткое (по отношению к конечностям), плоское и широкое туловище и, наконец, специфическое строение плечевого сочленения, обеспечивающее большую свободу вращения верхних конечностей в разных плоскостях. По-видимому, все перечисленные признаки были приобретены ранними гоминоидами в результате приспособления к таким способам передвижения по деревьям, которые требуют вертикального и хотя бы частично выпрямленного положения тела. Это лазание с опорой на нижние конечности, а также так называемая брахиация, т. е. перенос или перебрасывание тела с ветки на ветку с помощью верхних конечностей (рис. 6). Для низших обезьян ни то, ни другое, в общем, не свойственно, и они, в отличие от человекообразных, даже по ветвям передвигаются, как правило, на четырёх конечностях, подобно всем остальным млекопитающим от белки до леопарда. 
Одно время некоторые исследователи полагали, что церкопитекоиды и гоминоиды разделились ещё в раннем олигоцене, и что уже проплиопитеки и египтопитеки, жившие примерно 30–35 млн лет назад, должны рассматриваться как гоминоиды. Действительно, зубы этих обезьян, найденные в Файюмской впадине, несут хорошо выраженный узор дриопитека, но вот кости их черепа и скелета по своему строению ближе к аналогичным костям церкопитекоидов. Такая мозаичность признаков позволяет видеть в этих родах более или менее близкое подобие той предковой формы, от которой произошли церкопитекоиды и гоминоиды. К сожалению, огромный временной интервал, захватывающий весь поздний олигоцен, всё ещё остаётся практически неохарактеризованным ископаемым материалом, и поэтому представить себе сколько-нибудь детально процесс расхождения двух ветвей узконосых обезьян пока невозможно. Одно время в качестве наиболее ранней формы гоминоидов предположительно рассматривали род камояпитек (Kamoyapithecus), выделенный по находкам на позднеолигоценовом местонахождении Лосидок в северной Кении. Благодаря своему залеганию между двумя хорошо датированными калийаргоновым методом слоями базальта, нижний из которых имеет возраст 27,5±0,3 млн лет, а верхний 24,2±0,3 млн лет, эти находки имеют надёжную хронологическую привязку. Однако они всё же слишком малочисленны и фрагментарны, чтобы их с полной уверенностью можно было определить как останки человекообразной обезьяны. Более представительный материал, проливающий свет на ранние этапы эволюции гоминоидов, происходит с ряда местонахождений на западе Кении, но даже древнейшее из них — Месва Бридж — примерно на 3 млн лет моложе, чем Лосидок. Сейчас, благодаря находкам в Африке и Евразии, известно около 30 родов миоценовых гоминоидов, но предполагается, что этот материал и наполовину не отражает их действительного разнообразия. По некоторым оценкам, число родов, существовавших в названный период, могло быть в пять раз больше, и те из них, которые имеют критическое значение для понимания филогенетических взаимоотношений разных групп внутри надсемейства человекообразных, ещё не открыты.[3] Так это или нет, но представления о филогенезе гоминоидов — как ископаемых, так и современных — пока и в самом деле далеки от ясности. С середины 60-х гг. XX в. для построения генеалогического древа отряда приматов (а также многих других групп животных) стали использовать информацию, содержащуюся в макромолекулах белков и особенно нуклеиновых кислот. Принцип, лежащий в основе применяемых для этого методов, отчасти сродни тому, на котором базируются радиоизотопные способы датирования. Если в последних в качестве основы расчётов используется примерно одинаковая для больших промежутков времени скорость распада радиоактивных элементов (например, C14 — радиоактивного углерода), то в первых аналогичную роль играют так называемые нейтральные точечные мутации. Такие мутации, хотя и ведут к изменению последовательности нуклеотидов ДНК, не имеют, как предполагается, значения для естественного отбора и распределяются во времени (конечно, речь идёт о достаточно длительных его отрезках) более или менее равномерно. Если это так, то, сравнивая посредством различных, весьма изощрённых, методик строение молекул ДНК у разных групп организмов, можно судить о степени их родства (чем оно ближе, тем меньше должно быть различий), а при известной скорости мутирования даже и о примерном времени расхождения от общего предка. Конечно, биомолекулярные методы филогенетических исследований нельзя считать абсолютно надёжными и самодостаточными, и в этой области имеется ещё множество нерешённых проблем. Однако, как показывает опыт, в том, что касается эволюции приматов, биомолекулярный и палеонтологический анализ дают в общем, как правило, достаточно близкие результаты. Сопоставление последовательностей нуклеотидов в молекулах ДНК, взятых у современных церкопитековых и человекообразных обезьян, говорит, по мнению большинства специалистов, о том, что эволюционные пути этих групп разошлись где-то в интервале от 22 до 28 млн лет назад. Таким образом, палеонтологические и молекулярные данные, взятые вместе, позволяют считать, что самостоятельная филогенетическая история надсемейства гоминоидов, в состав которого из ныне живущих приматов входят человек и человекообразные обезьяны (шимпанзе, горилла, орангутанг, гиббон, сиаманг), началась около 25 млн лет назад (рис. 4). До недавнего времени в составе надсемейства гоминоидов было принято выделять три семейства: гилобатид (Hylobatidae), представленных гиббоном и сиамангом, понгид (Pongidae), к которым относили роды орангутанга (Pongo), гориллы (Gorilla) и шимпанзе (Pan), и гоминид (Hominidae), т. е. человека и его прямоходящих предков. Эта классификация основывалась на внешних анатомических признаках, прежде всего таких, как пропорции конечностей, особенности строения клыков и коренных зубов и т. п. Широкое использование биомолекулярных методов в систематике, однако, показало, что необходима перегруппировка принятых к настоящему времени таксонов. В частности, выяснилось, что орангутанг в генетическом плане отстоит от африканских человекообразных обезьян (горилла и шимпанзе) дальше, чем последние от человека, и должен быть выделен в особое семейство. Кроме того, появились данные, говорящие о том, что генетическая дистанция между человеком и шимпанзе может быть даже меньше, чем между шимпанзе и гориллой, и, если это так, то, значит, необходимы и соответствующие изменения в систематике. Практически нет сомнений в том, что появились гоминоиды в Африке, и почти 10 млн лет их история оставалась связанной исключительно с этим континентом. Если не считать упоминавшиеся выше спорные материалы из Лосидок, то самые ранние гоминоиды, найденные на нижнемиоценовых местонахождениях Восточной Африки, относятся к роду проконсул (Proconsul) (рис. 7). Правда, существует точка зрения, согласно которой проконсул тоже не являлся ещё собственно гоминоидом, но и её сторонники допускают, что какой-то из видов этого рода вполне мог быть общим предком всех позднейших человекообразных обезьян. 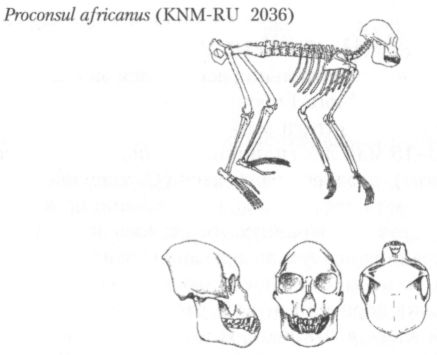
В конце раннего миоцена в Африке жили уже представители нескольких родов гоминоидов: дендропитеки, микропитеки, афропитеки, турканопитеки и др., но филогенетическое значение этих форм неясно. Трудно сказать, имела ли какая-то из них непосредственное отношение к родословной современных горилл или шимпанзе. По размеру тела африканские раннемиоценовые гоминоиды варьировали от очень мелких, до 3 кг весом (Micropithecus clarki), до крупных (Proconsul major, Turkanapithecus heseloni), весящих около 100 кг, как самка современной гориллы, а их рацион включал главным образом фрукты и молодые листья. Все перечисленные формы вели преимущественно древесный образ жизни, а при передвижении по земле оставались четвероногими.[4] Единственным исключением из последнего правила был, возможно, ореопитек, или, точнее, вид Oreopithecus bamboli, но он обитал не в Африке, а в Европе, и не в начале, а в конце миоцена. Изучение костных останков ореопитека, найденных в Италии в отложениях возрастом 8–9 млн лет, привело ряд палеонтологов к предположению, что это существо, когда оно оказывалось на земле, предпочитало использовать для ходьбы не четыре, а две ноги. В среднем миоцене, когда между Африкой и Евразией установилась сухопутная перемычка, (16–17 млн лет назад), ареал обитания гоминоидов значительно расширился за счёт включения в него территорий юга Европы и Азии. Наиболее древние ископаемые представители этой группы в Европе имеют возраст около 13–15 млн лет (плиопитек (Pliopithecus), дриопитек (Dryopithecus), позднее уранопитек (Ouranopithecus)), а в Азии порядка 12 млн лет. Однако, если в Азии, по крайней мере на её юго-восточных окраинах, им удалось основательно закрепиться, сохранившись там до сего дня (орангутанги, гиббоны, сиаманги), то в Европе условия оказались менее подходящими, и, пережив короткий период расцвета, к концу миоцена гоминоиды здесь вымирают. В отложениях возрастом моложе 7 млн лет их останков в Европе не найдено. В Африке в рассматриваемый период времени (от 15 до 5 млн лет назад) также отмечается значительное сокращение числа известных видов гоминоидов, но, несмотря на это, именно она всё же остаётся местом основных событий в их эволюции. О важнейших из этих событий, непосредственно связанных с происхождением человека, и пойдёт речь в следующих главах. Примечания:[1] См., например: Татаринов Л. П. Очерки по теории эволюции. М., 1987. С. 186–188; Будыко М. И. Путешествие во времени. М., 1990. С. 16. [2] По-гречески «нос» — ???, слово «?????» — форма этого существительного в родительном падеже. (Примеч. ред.) [3] Ward C. V. et al. Function and phylogeny in Miocene hominoids // Function, Phylogeny, and Fossils: Miocene Hominoid Evolution and Adaptations. New York, 1997. P. 1–2. [4] Pilbeam D. Research on Miocene hominoids and hominid origins. The last three decades // Function, Phylogeny, and Fossils: Miocene Hominoid Evolution and Adaptations. New York, 1997. |
|
||
|
Главная | Контакты | Нашёл ошибку | Прислать материал | Добавить в избранное |
||||
|
|
||||