|
||||
|
|
Глава 4. За гранью: род Homo Трудные времена Мы подошли к очень важному, можно сказать, решающему моменту в эволюции наших предков, моменту, когда вследствие удачного соединения в одном месте и в одно время ряда естественных факторов этот процесс приобрёл совершенно специфический характер и необычное направление. Те гоминиды, о которых речь шла до сих пор, оставались, несмотря на формальную принадлежность к кладе человека, по сути всё-таки ещё животными — в их поведении не было ничего такого, что сильно выделяло бы их среди других обитателей африканских джунглей и саванн. Конечно, в отличие от остальных обезьян, они передвигались по земле, образно говоря, с высоко поднятой головой, используя не четыре, а только две конечности, но так же поступают, например, и страусы, да и среди млекопитающих любители походить на двух ногах тоже встречаются (были они и среди динозавров). Что же касается способов добывания пищи, защиты от врагов, выращивания потомства, т. е., иными словами, характера приспособления к среде, то здесь между первыми гоминидами и их четвероногими собратьями практически никаких различий не было. Такие различия становятся более или менее заметны лишь начиная с конца плиоцена (2,5–1,7 млн лет назад), когда, судя по археологическим данным, появляются самые древние следы существования того, что называется культурой. Как уже говорилось в первой главе, единственное существенное отличие эволюции гоминид от эволюции всех других живых существ заключается в том, что нашим предкам приходилось приспосабливаться не только, а с известного момента и не столько к естественной среде, сколько к среде искусственной, культурной. Этого отличия оказалось вполне достаточно, чтобы антропогенез приобрёл характер более чем специфический для чисто биологического процесса и привёл к появлению тех многочисленных поведенческих и анатомических особенностей, которые столь явно выделяют людей даже на фоне их ближайших родственников — человекообразных обезьян. Культура — центральное понятие для наук о человеке вообще — является, таким образом, ключевым и при объяснении не совсем обычного характера нашей биологической эволюции. Взятое в самом широком смысле, это понятие означает особую сферу бытия, в известном смысле противостоящую природе. Её существование возможно лишь при условии появления у живых существ такого поведения, в основе которого лежит не программа, заложенная в генах, а информация, накапливаемая посредством научения, т. е. благодаря способности к запоминанию индивидуального опыта и его пополнению через наблюдение, подражание, умозаключение и т. д. Однако это условие хоть и необходимое, но ещё не достаточное, поскольку, скажем, условный рефлекс — тоже результат научения, внегенетического усвоения информации, но вряд ли кто-то станет считать культурным феноменом выделение желудочного сока у подопытного животного в ответ на привычный раздражитель. Поэтому в наше определение нужно ввести ещё один критерий. Этот критерий — избирательность. Поведение, связанное с культурой, формируется не только надгенетически, но и не механически, т. е. активно, избирательно. Появление и развитие такого поведения означает рост количества свободы в мире. Пожалуй, не будет большим преувеличением утверждать, что оно возвещает конец эры детерминизма и предсказуемости и начало эры свободы и непредсказуемости (простой пример: поведение большинства животных в той или иной конкретной ситуации «просчитать» гораздо проще, чем поведение большинства людей). Подытоживая, можно сформулировать такое определение: культура — это все формы поведения, основанные на внегенетически и притом избирательно (не механически) усваиваемой, хранимой и передаваемой информации, а также их результаты (вещественные и идеальные). Благодаря исследованиям приматологов и зоопсихологов, сейчас можно, пожалуй, уже не сомневаться в том, что интеллектуальные возможности шимпанзе, орангутангов и ряда других обезьян вполне достаточны для осуществления ими весьма сложных форм культурного поведения. Информация, приобретаемая через научение, играет в их жизни едва ли не большую роль, чем врождённая, генетическая программа. Не удивительно, что в последние годы, в связи с обнаружением всё новых и новых доказательств культурной «продвинутости» высших приматов, на стыке приматологии, зоопсихологии и антропологии стала формироваться особая дисциплина, не имеющая пока ещё общепринятого наименования, но, по сути, представляющая собой не что иное, как культурную приматологию.[22] Вполне вероятно, что уже через несколько лет это словосочетание станет для нас таким же привычным, каким сегодня является культурная антропология. Человекообразные обезьяны способны изготавливать орудия, общаться между собой и с людьми посредством выученных знаков, они умеют даже лгать, и, скорее всего, обладают самосознанием и рядом других качеств, которые долгое время считались исключительно человеческими. Однако вместе с тем, в естественных условиях поведенческий репертуар даже самых развитых приматов включает лишь отдельные элементы культуры, не связанные в сколько-нибудь целостную, имеющую жизненно важное значение систему. Лишь у гоминид выживание, начиная с определённого момента, стало прямо зависеть от «вовлечённости» в культуру, и именно эта зависимость породила в своё время, по выражению американского биолога Э. Майра, «совершенно новые давления отбора»[23], результатом чего и стало появление столь необычного существа, как человек. Почему же это произошло, почему роль культуры в жизни наших предков столь несоизмеримо возросла по сравнению с её ролью в жизни других гоминоидов, не говоря уж об остальных приматах? Вероятнее всего, это произошло потому, что на определённом этапе своей истории гоминиды, или по крайней мере какая-то их часть, один или несколько видов, оказались в такой ситуации, когда приспособления к меняющейся природной среде обычным биологическим путём оказалось недостаточно для выживания, и единственным выходом стало использование скрытых, невостребованных дотоле резервов интеллекта. Самые ранние гоминиды — те, которые жили в конце миоцена — начале плиоцена, т. е. в период примерно от 6 до 3 млн лет назад — вероятно, лишь немногим отличались по своему поведению от современных шимпанзе, да и от других крупных человекообразных обезьян. Конечно, вполне возможно, что они чаще прибегали к помощи разного рода орудий, обладали повышенной, по сравнению со всеми другими приматами, социальностью и т. п., но серьёзных оснований предполагать, что культура играла в их жизни сколько-нибудь заметно возросшую роль, нет. Их мозг, как уже говорилось в предыдущей главе, по размерам и строению был ещё вполне обезьяньим, и таким же оставалось их существование в целом. Это и неудивительно, поскольку первые несколько миллионов лет своей истории австралопитеки, не говоря уже об их предшественниках, имели ещё возможность вести частично древесный образ жизни и сохраняли необходимые для этого анатомические приспособления, что позволяло им чувствовать себя достаточно комфортно в естественном состоянии. Как показало изучение древних почв и ископаемой пыльцы растений с местонахождений, откуда происходят кости австралопитеков, ландшафты, в которых они обитали, в большинстве случаев были закрытыми (лес) либо смешанными. Ещё более древний Ardipithecus ramidus kadabba также был, судя по результатам проведённых в Среднем Аваше палеогеографических исследований, жителем влажного леса. Однако климат, как мы уже знаем, менялся, и в конце плиоцена, примерно 2,5–3 млн лет назад, для гоминид наступили очень трудные времена. Само их существование оказалось под угрозой. Начался очередной цикл иссушения климата, и природные условия восточных районов Африки претерпели резкие изменения. Стало значительно холоднее, чем прежде, сильно сократилось количество осадков. В результате площади, занимаемые тропическими лесами, существенно сократились, а саванны и близкие им ландшафты открытого и полуоткрытого типа, наоборот, получили большое распространение. Климатические и ландшафтные перемены не могли не отразиться на состоянии животного мира: необходимость приспособления к новым условиям вызвала в рассматриваемый период настоящий взрыв видообразования среди самых разных групп млекопитающих. Для интервала от 3 до 2,5 млн лет назад зафиксировано появление множества (десятков) новых видов крупных копытных животных и грызунов, а также вымирание ряда старых таксонов. Аналогичные процессы происходили и среди гоминид. В частности, можно вспомнить, что именно в это время прекратил своё существование вид австралопитек афарский (Australopithecus afarensis), и на смену ему пришло несколько форм грацильных и массивных австралопитеков: австралопитек африканский (Australopithecus africanus), австралопитек гархи (Australopithecus garhi), австралопитек эфиопский (Australopithecus aethiopicus), (см. табл. 1 и рис. 10). Появление первых представителей рода Homo, датируемое чуть более поздним периодом (2,3–2,4 млн лет назад), также происходило на фоне продолжавшегося иссушения климата и сопряжённых с ним ландшафтных перестроек. Вероятно, это был самый критический эпизод во всей истории древнейших предков человека, когда они буквально балансировали на грани вымирания. О том, сколь трудным было положение, в котором ранние гоминиды с их несовершенной ещё двуногостью и недостаточно развитыми органами защиты и нападения оказались в результате описанных природных пертурбаций, свидетельствуют данные по их демографии. Речь идёт, конечно, не о численности популяций — здесь сколько-нибудь точные оценки вряд ли возможны, — а об изменениях продолжительности жизни представителей разных видов. Дело в том, что в ряде случаев ископаемые кости позволяют более или менее точно установить, на какой стадии онтогенеза (т. е. индивидуального развития) прервалась жизнь особи, которой они принадлежали. Об этом судят, в частности, по степени прорезанности или стёртости зубов, по состоянию швов, разделяющих кости черепа, по срощенности различных сегментов длинных костей и т. д. Определить возраст в годах можно далеко не всегда, тем более что темпы онтогенеза за миллионы лет эволюции тоже претерпевали изменения, но зато часто удаётся сделать достаточно обоснованное заключение о том, к какой возрастной группе принадлежала данная особь, т. е. имеем ли мы дело, например, с останками ребёнка, зрелого индивида или старика. Изученные таким образом костные материалы говорят о том, что на протяжении интересующего нас критического периода процент гоминид, погибавших или умиравших естественной смертью, не достигнув зрелого возраста, постоянно возрастал, став в итоге просто устрашающим. Если из 63 образцов костей австралопитека африканского, допускающих возрастные определения, лишь 35% принадлежали, по подсчётам Ф. Тобайаса, особям, не достигшим зрелости, то для 119 таких образцов, относимых к более позднему виду австралопитека массивного (Australopithecus robustus), этот показатель, согласно определениям А. Манна, составляет 60,5%. Наконец, для ещё более позднего хабилиса (Homo habilis) из Олдувайского ущелья (22 образца) он, по данным Тобайаса, достигает уже 73%.[24] Разумеется, приведённые цифры отражают действительную динамику смертности в популяциях поздних австралопитеков и ранних Homo лишь приблизительно, и при изменении величины выборок в итоговую картину могут быть внесены существенные коррективы, но общая тенденция вырисовывается всё же слишком отчётливо, чтобы можно было поставить её под сомнение. Следует, к тому же, иметь в виду, что все указанные здесь процентные показатели скорее занижены, чем завышены по сравнению с реальными, поскольку кости молодых особей — особенно детские — сохраняются в ископаемом состоянии гораздо хуже, чем кости особей взрослых. Описанное изменение доступных для оценки демографических параметров (кстати, прямо свидетельствующее об интенсификации работы естественного отбора), несомненно, было обусловлено разрушением привычной среды обитания и недостаточной физической приспособленностью гоминид к новым условиям. Всё это вполне могло бы иметь для них — а для ряда видов и в самом деле имело — фатальные последствия, но у обострявшейся кризисной ситуации была и оборотная сторона. Поставив гоминид на грань вымирания, кризис в то же время стимулировал развитие культурного поведения, первые осязаемые свидетельства которого не случайно фиксируются как раз тогда, когда вопрос «быть или не быть» должен был приобрести применительно к судьбе человечества своё буквальное значение. Именно к этому периоду относятся древнейшие достоверные каменные орудия (они имеют возраст порядка 2,5–2,6 млн лет), и именно этим периодом следует, по-видимому, датировать начальную стадию процесса, который можно определить как первую культурную революцию в истории человечества. Суть её заключалась во «втягивании» наших предков в культуру, ставшую в итоге для них главным и необходимым средством приспособления к естественной среде. Разумеется, первая культурная революция была революцией не в смысле своей быстроты — она растянулась, как минимум на сотни тысяч лет — а по своим последствиям, по тем изменениям, которые произошли в поведении гоминид, и по тому влиянию, которое она оказала на весь ход их дальнейшей истории. Главным итогом этой революции стало превращение культуры в фактор, определяющий и организующий все основные аспекты жизнедеятельности гоминид, а также и их биологического развития. Нельзя не заметить, сколь большую роль в определении направления нашей эволюции сыграл случай, буквально вынудивший плиоценовых гоминид «вспомнить» о культуре. В самом деле, ведь случаем мы называем совпадение событий, которые не находятся между собой в отношении причины и следствия и не зависят от одной общей причины, т. е. событий, между которыми не существует никакой причинной связи, а «пусковой механизм» антропогенеза был «включён» как раз благодаря такому совпадению. Толчком, заставившим его начать работу, послужило маловероятное в принципе пересечение в нужном месте и в нужное время практически независимых друг от друга биологических (формирование у какой-то группы или групп интеллектуально продвинутых гоминоидов предрасположенности к прямохождению), климатических (аридизация) и тектонических (образование Восточноафриканского рифта) процессов. Это случайное стечение обстоятельств закрыло для наших предков возможность адаптации к меняющимся условиям существования обычным — биологическим — путём и подтолкнуло к более активной реализации уже имевшегося у них достаточно высокого интеллектуального потенциала. А что было бы, если бы обстоятельства сложились иначе? Этот вопрос сейчас очень любят задавать историки, изучающие сравнительно недавнее прошлое, но по отношению к первобытной истории и, в частности, к эволюции человека он имеет не меньшую важность. Итак, что было бы, если бы, скажем, к моменту пика аридизации среди гоминоидов Восточной Африки не оказалось двуногих «монстров»? Или, наоборот, если бы они были, а аридизация и смена ландшафтов так и не начались? Или если бы Восточная Африка не оказалась отделённой от остальной части материка рифтовой системой? С абсолютной точностью просчитать все эти варианты, конечно, невозможно, но, скорее всего, в каждом из этих случаев обезьяны ещё на миллионы лет (а то и навсегда) остались бы обезьянами, и не исключено, что рано или поздно вместо них на «магистральный» путь эволюции вышли бы какие-то иные животные. Основатели рода Австралопитеки исчезли с лица земли только в плейстоцене (см. табл. 3 и табл. 4), успев задолго до этого дать начало роду Homo и будучи, следовательно, свидетелями, а в какой-то степени, по-видимому, и жертвами эволюционного триумфа новой формы гоминид. Дольше других просуществовали «массивные» австралопитеки, т. е. парантропы, самые поздние представители которых жили ещё полтора миллиона или даже миллион лет назад. Их окончательное исчезновение в нижнем плейстоцене одни исследователи объясняют неблагоприятными климатическими изменениями, другие — конкуренцией со стороны более развитых гоминид, а третьи, как это ни странно, вымиранием саблезубых тигров. В последнем случае предполагается, что тигры, пока они ещё жили в Африке, хотя и охотились на австралопитеков, но в то же время ограждали их от других хищников, гораздо более многочисленных. Когда же этой защиты не стало, то львы, леопарды и гиены быстро разделались с и без того уже вырождавшимися парантропами, сильно отставшими в своём развитии от других гоминид. Среди надёжно датированных ископаемых материалов, относимых к роду Homo, наибольшую древность — 2,4 млн лет — имеет височная кость с местонахождения Туген Хиллс в Кении. Она происходит из отложений формации Чемерон. Правда, установить не только родовую, но и видовую принадлежность находки в данном случае очень трудно ввиду её фрагментарности. Вообще же первым представителем рода Homo, или промежуточным звеном между ним и австралопитеками, считается, с рядом оговорок, вид хабилис (Homo habilis — на латыни это означает «человек умелый»). Этот вид обособился, вероятно, около 2,4 млн лет назад. Интересно, что примерно к этому же времени относятся и древнейшие из известных сейчас каменных орудий со следами намеренного изготовления, которые знаменуют собой для археологов начало палеолита (древнекаменного века) и начало нашей культурной истории в целом. Самая ранняя стадия этой истории получила название олдувайской эпохи. Оно происходит от наименования ущелья Олдувай в Танзании, где английский археолог Луис Лики в конце 50-х — начале 60-х гг. прошлого века впервые обнаружил грубые изделия из кусков лавы и кварцевых галек (рис. 17), изготовленные то ли кем-то из поздних австралопитеков, то ли первыми представителями рода Homo.
 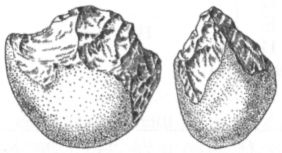
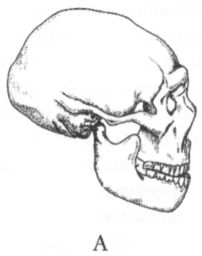
Хотя Homo в переводе с латыни означает «человек», из этого широко известного факта не следует делать вывод, что человек и Homo — одно и тоже. «Человек», «люди» — это понятия совсем другого смыслового ряда, нежели понятия «гоминиды» или «члены рода Homo». Они подразумевают совершенно разные классификации живых существ, и поэтому не следует воспринимать их как синонимы. В первом случае мы имеем дело с философской классификацией, а во втором — с биологической. Люди, человек — это существа, специализированные к культуре, существа, для которых культура является и программой поведения, и средством адаптации, и средой («встроенной») обитания. Культура — это то, что делает Homo, потенциальных людей, людьми действительными. Для того чтобы быть членом вида Homo sapiens, или карликовый шимпанзе (Pan paniscus), или волк (Canis lupus) достаточно просто родиться, а для того, чтобы быть человеком, надо им стать, т. е. подвергнуться воздействию искусственной среды, культуры. В этом смысле можно сказать, что качественная разница между животными и людьми состоит в том, что первыми рождаются, а вторыми становятся, становятся лишь некоторое время спустя после рождения и лишь при наличии определённых условий (т. е. необходимых биологических предпосылок и культурной среды). Поэтому, кстати, вряд ли стоит искать, как это часто делали и иногда всё ещё делают, некие анатомические критерии (вроде «мозгового Рубикона»), якобы разрешающие пресловутую «проблему грани». Во всяком случае, не стоит придавать такого рода критериям абсолютное значение. С одной стороны, вполне вероятно, что уже самые ранние представители рода Homo в большинстве своём подпадали под определение «человек» (к сожалению, мы слишком мало знаем об их поведении, чтобы утверждать это с уверенностью). С другой стороны, даже среди членов вида Homo sapiens могут изредка встречаться особи, в силу неустранимых анатомических патологий или несчастных превратностей биографии не испытавшие должного влияния культурной среды. Костные останки хабилиса — два зуба — были впервые обнаружены экспедицией Лики в Олдувайском ущелье в 1959 г. Поначалу они не привлекли особого внимания, поскольку в том же полевом сезоне неподалёку от них был найден почти целый череп массивного австралопитека, вошедший в историю науки под именем «зинджантропа». В относительной близости от него залегали примитивные каменные орудия, творцом которых и был поначалу признан зинджантроп. Однако уже в следующем году на несколько большей глубине был найден ещё один череп, своим строением гораздо сильнее напоминавший человеческий и, главное, заметно превосходивший череп зинджантропа по объёму мозговой полости. Тогда вспомнили и о зубах, обнаруженных ранее, и выделили на основе этих материалов вид Homo habilis. Статья, возвещавшая об этом, была опубликована в 1964 г., но широкое признание новая группа гоминид получила лишь в конце 70-х — начале 80-х гг. Произошло это во многом благодаря открытию в Кооби-Фора на восточном берегу озера Туркана (Кения) двух хорошо сохранившихся черепов, по своему строению занимавших явно промежуточное положение между австралопитеками, с одной стороны, и более поздними питекантропами (Pithecanthropus или Homo erectus), с другой. Кроме Олдувая и оз. Туркана (в старой литературе его называют оз. Рудольф), скелетные материалы, относимые к H. habilis, были обнаружены ещё в Хадаре (Эфиопия), в Урахе (Малави) и, предположительно, в южноафриканских в пещерах Штеркфонтейн и Сварткранс. Если кости конечностей хабилиса, в общем, довольно мало отличаются от аналогичных участков скелета австралопитеков, а по пропорциям могут быть даже близки к архаичному типу, установленному для австралопитека африканского, то зубы и особенно череп характеризуются множеством новых важных черт строения. Менее резким становится выступание нижней части лица, не столь заметна массивность нижней челюсти, зубная дуга приобретает форму, более близкую к параболической, а коренные зубы в некоторых случаях уменьшаются в размере и эмаль на них утончается. Свод черепа становится несколько выше, затылочная часть делается более округлой (хотя и сохраняет ещё угловатость), а главное, ряд серьёзных изменений претерпевает мозг. Наиболее существенные из этих изменений заключались в увеличении его абсолютного и относительного размера, значительном расширении теменной области, формировании подобного человеческому узора борозд и появлении достаточно хорошо выраженных специфических выпуклостей в областях, где у людей находятся речевые центры (поле Брока? и поле Вернике). По мнению авторитетного южноафриканского антрополога Ф. Тобайаса, именно с появлением хабилиса (Homo habilis) в эволюции мозга гоминид «был сделан гигантский шаг к новому уровню организации».[25] Действительно, даже если судить только по величине мозга, не касаясь его строения, то различия между австралопитеками и человеком умелым всё равно будут впечатляющими. У первых объём эндокрана колеблется от 400 до 500 см3, почти никогда не выходя за эти рамки, а у второго он редко бывает меньше 600 см3 и в отдельных случаях достигает даже 700 с лишним кубических сантиметров. Вопрос о том, существует ли прямая связь между размером мозга и степенью развития интеллекта, является дискуссионным. Одни специалисты считают, что такой связи нет. Как заявил по этому поводу американский исследователь Г. Джепсер: «Мозговая полость подобна кошельку, содержимое которого значит гораздо больше, чем его размер». Другие, напротив, уверены в том, что связь есть, и что в целом существует стойкая положительная корреляция между размером головного мозга, с одной стороны, и коэффициентом интеллектуального развития, с другой. Так это или нет, но, что касается прогрессирующего увеличения мозга у членов рода Homo, то кажется почти несомненным, что в качестве главного фактора, обусловившего этот процесс, выступала именно возраставшая роль интеллекта и культуры. Эта уверенность основана не только на том обстоятельстве, что первый заметный скачок в размере эндокрана у гоминид (хабилис) хронологически совпадает с появлением древнейших каменных орудий и других археологических свидетельств усложнения культурного поведения. Дело ещё и в том, что мозг, наряду с сердцем, печенью, почками и кишечником, входит в число наиболее «дорогостоящих» в энергетическом отношении анатомических органов. В то время как общий вес перечисленных органов составляет у человека в среднем всего лишь 7% от веса тела, потребляемая ими доля метаболической энергии превышает 75%. Вес мозга составляет 2% от веса тела, потребляет же он примерно 20% получаемой организмом энергии. Чем больше мозг, тем больше сил и времени приходится его обладателю тратить на добывание пищи. Вместо того, чтобы спокойно отдыхать в укромном месте, он вынужден лишние часы бродить по джунглям или саванне в поисках съедобных растений и животных, ежеминутно рискуя при этом превратиться из охотника в жертву более сильных хищников. Поэтому для большинства видов крупный мозг, такой, как у приматов и особенно человека — непозволительная роскошь. Увеличение его размера могло стать возможным лишь в том случае, если сопутствующий этому процессу рост энергетической нагрузки на организм компенсировался какими-то важными преимуществами, обеспечивавшими благоприятное для «высоколобых» действие естественного отбора. Учитывая же функции мозга, трудно сомневаться в том, что преимущества эти были связаны в первую очередь именно с развитием интеллекта (памяти, мыслительных способностей) и соответствующими изменениями в поведении. В этом отношении не случайным, по-видимому, является ещё одно хронологическое совпадение. Археологические данные дают основания полагать, что появлению рода Homo сопутствовали изменения в характере питания предков человека, а именно, увеличение потребления мяса. Хотя характер стёртости зубов у гоминид олдувайской эпохи говорит о том, что основу их рациона всё ещё составляли растительные продукты, мясная пища, как видно из обилия костей на некоторых древнейших стоянках, а также из наличия орудий, служивших для разделки туш, тоже приобрела уже немаловажное значение. Это можно считать важным условием роста мозга, поскольку сокращение доли растительной пищи в рационе наших предков и рост доли пищи животной — более калорийной и легче усваиваемой — создавали возможность для уменьшения размеров кишечника, который, как уже говорилось, также входит в число энергетически наиболее дорогостоящих органов. Такое уменьшение должно было помочь поддержать общий метаболический баланс на прежнем уровне, несмотря на значительный рост мозга. Не случайно у современного человека кишечник много меньше, чем у других животных сходного размера, причём получаемый за счёт этого энергетический выигрыш обратно пропорционален потерям, связанным с увеличившимся мозгом. Давно уже было замечено, что материалы, относимые к Homo habilis, — а в основном это фрагменты черепов — довольно сильно различаются между собой. Свойственная им высокая степень различий явно превышает таковую у современных людей и может объясняться по-разному. Одни исследователи считают её результатом постепенной эволюции в пределах вида, обосновывая эту точку зрения тем, что самые древние и самые молодые находки разделяет промежуток времени в несколько сотен тысяч лет. Другие также полагают, что наблюдаемая изменчивость является внутривидовой, но объясняют её развитым половым диморфизмом, т. е. большими различиями между самцами и самками Homo habilis (может быть, в данном случае уместней уже было бы сказать «между мужчинами и женщинами») по размерам и некоторым особенностям строения скелета. Наконец, согласно третьей точке зрения, приобретшей особенную популярность в последнее десятилетие и являющейся сейчас преобладающей, существующую вариабельность следует рассматривать как отражение таксономической неоднородности группы. Многие антропологи стали проводить различие между Homo habilis sensu lato (хабилис в широком смысле) и Homo habilis sensu stricto (т. е. хабилис в узком смысле или собственно хабилис), и те находки, которые не могут быть отнесены к последнему таксону, зачислять в вид человек рудольфский (Homo rudolfensis) (рис. 18). Допускается также, что южноафриканские образцы могут представлять ещё один, третий, вид, не известный в Восточной Африке, но общепринятого названия для него пока не предложено. 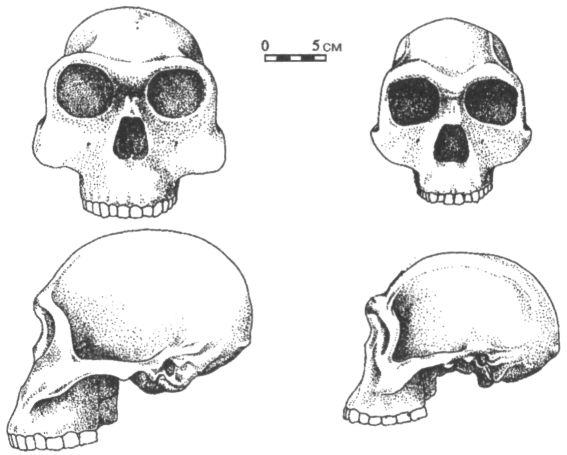
Главную роль в выделении антропологами вида человека рудольфского (Homo rudolfensis) сыграл череп за инвентарным номером 1470, найденный в 1972 г. на местонахождении Кооби-Фора в Кении. Он отличается крупными размерами и слишком большим даже для хабилисов объёмом мозговой полости (750 см3). Ознакомившись с его описанием, российский исследователь В. П. Алексеев назвал обладателя этого черепа «питекантропом рудольфским» — Pithecanthropus rudolfensis (Кооби-Фора находится на берегу озера Туркана, которое, напомню, в колониальные времена было известно как озеро Рудольф). Впоследствии, поскольку родовое имя питекантроп (Pithecanthropus) практически вышло из употребления, оно было заменено на Homo, а вот видовое название, предложенное Алексеевым, прижилось и широко используется в специальной литературе. Сейчас иногда ставится под сомнение не только единство хабилисов как вида, но даже и правомерность отнесения их к роду Homo. Для этого есть некоторые основания. По многим анатомическим характеристикам, которые могут быть более или менее надёжно реконструированы и которые тесно связаны с экологическими, демографическими, репродуктивными и прочими особенностями поведения вида, хабилисы, действительно, явно ближе к австралопитекам, чем к более поздним гоминидам — членам рода Homo. Среди этих характеристик такие, как размер и форма тела, пропорции конечностей, темпы индивидуального развития, особенности жевательного аппарата и т. д. Правда, при сравнении по размеру мозга картина получается не столь однозначной, и всё же некоторые антропологи считают, что правильнее в настоящее время переместить виды хабилиса и человека рудольфского в род австралопитеков. После открытия черепа из Ломекви, послужившего для выделения рода кениантроп (Kenyanthropus) и оказавшегося довольно похожим по строению лицевой части на череп 1470, было высказано также предположение, что человек рудольфский мог представлять собой более поздний, чем platyops, вид кениантропа. В этом случае его правильное научное название — Kenyanthropus rudolfensis. Судя по имеющимся сейчас данным, хабилисы (в широком смысле) жили примерно от 2,4 до 1,7 млн лет назад, после чего им на смену пришли более «продвинутые» гоминиды. В современной литературе часто высказывается точка зрения, что предком последних мог быть человек рудольфский, но точно сказать, какая из двух (или трёх?) форм Homo habilis sensu lato ближе стоит к главному стволу эволюции человека, пока невозможно. В любом случае, однако, иных вариантов, кроме как выводить позднейших Homo из хабилисов, в настоящее время просто не существует. «Массивные» австралопитеки, заблудившиеся в лабиринте эволюции и кончившие свои дни в его глухих тупиках, на роль наших прямых предков претендовать не могут, а другие ископаемые подходящего возраста пока отсутствуют. Да и сам хабилис как возможный прародитель, в конце концов, не так уж плох, независимо от того, будем ли мы считать его ещё австралопитеком или уже Homo. Архантропы Примерно 1,7 млн лет назад начался плейстоцен, или, иначе говоря, ледниковый период. Значительную часть Северного полушария к этому времени уже покрывали льды, а на примыкающих к ним пространствах господствовали тундры и лесотундры. В плейстоцене средняя температура на Земле постепенно понижалась, климат в целом становился хуже, но время от времени этот процесс приостанавливался, и во многих регионах суровые условия вновь сменялись относительно мягкими. В наиболее холодные периоды ледниковые языки и сопутствующие им ландшафты продвигались далеко на юг (почти до 50° с. ш.), а в эпохи потеплений — их называют межледниковьями — опять отступали к северу. Мы живём как раз в одно из таких межледниковий, которое, однако, принято выделять как самостоятельную геологическую эпоху, именуемую голоценом и не включаемую в плейстоцен. Таблица 4 показывает два основных варианта периодизации плейстоцена. В дальнейшем в книге будет использоваться только вариант А. Последние полтора с небольшим миллиона лет эволюции человека, т. е. как раз плейстоценовый её отрезок, можно с большей или меньшей степенью условности разделить на три частично перекрывающихся периода или стадии: стадию архантропов, стадию палеоантропов и стадию неоантропов. Архантропы — это общее название для гоминид, в прошлом именовавшихся питекантропами и синантропами, а сейчас относимых большинством антропологов к виду эректус (Homo erectus), который пришёл на смену хабилисам. Иногда этот вид делят на два и более раннюю форму называют эргастер (Homo ergaster). Термин палеоантропы может служить в качестве собирательного обозначения всех близких и дальних потомков архантропов, за исключением людей современного физического типа. Как правило, этих потомков зачисляют в виды человек гейдельбергский (Homo heidelbergensis) и неандерталец (Homo neanderthalensis), хотя есть и иные варианты их классификации. Наконец, неоантропы — это люди современного физического типа, т. е. члены вида гомо сапиенс (Homo sapiens). Правда, начиная с середины 60-х гг., многие исследователи стали обозначать людей современного физического типа как Homo sapiens sapiens, отводя им тем самым подвидовой ранг в пределах одного с неандертальцами (Homo sapiens neanderthalensis) или всеми палеоантропами вообще вида, но у такого подхода, кажется, больше недостатков, чем преимуществ. Главный из этих недостатков заключается в том, что расширительный подход к определению состава вида гомо сапиенс выглядит довольно спорно в свете приобретающих сейчас всё больший размах палеогенетических исследований. Сопоставление ДНК, извлечённой из костей классических неандертальцев, с ДНК современных людей показало, что генетическое расстояние между ними в три с лишним раза превышает расстояние между ныне существующими расовыми группами человека. Иными словами, если у современных негроидов, европеоидов и монголоидов идентичные фрагменты цепи ДНК (изучались так называемые гипервариабельные участки I и II), состоящие из 600 нуклеотидов, различаются между собой в среднем по 10 позициям, то при сравнении каждой из этих групп с неандертальцами число расхождений возрастает до 35.[26] По строению скелета и особенно черепа виды гомо сапиенс и Homo neanderthalensis тоже различаются между собой намного больше, чем следовало бы ожидать от представителей одного биологического вида. Таким образом, в данном случае лучше следовать традиционному словоупотреблению, оставив обозначение гомо сапиенс (Homo sapiens) лишь за людьми современного анатомического типа (что, разумеется, не следует понимать как отказ в «разумности» иным типам гоминид). Архантропы, судя по находкам на восточном (местонахождение Кооби-Фора) и западном (местонахождение Нариокотоме) побережьях озера Туркана, появились около 1,8 млн лет назад. Наиболее древних африканских представителей этой формы, отличающихся от остальных некоторыми особенностями строения мозгового и лицевого отделов черепа, часто выделяют сейчас как вид эргастер (Homo ergaster). Первоначально этот таксон был выделен по единственной челюсти, а названием своим, означающим в несколько вольном переводе с греческого «трудяга» или «работяга», он обязан тому обстоятельству, что поблизости от этой челюсти залегали каменные орудия. Иногда правомерность обособления ранней формы архантропов в качестве отдельного вида оспаривается. При этом указывают, например, на то, что вариабельность черепов представителей рода Homo возрастом от 1,7 до 0,5 млн лет не превышает таковой внутри современного человечества, представляющего собой один биологический вид. По этой и по ряду других причин некоторые антропологи считают, что выделение в составе архантропов особого вида эргастер недостаточно обосновано, и предпочитают по-прежнему говорить о едином виде эректус (Homo erectus). Кроме того, недавно было высказано предложение выделять ранних представителей этого вида как хронологический подвид, именуя его Homo erectus ergaster. Первые полмиллиона лет своего существования архантропы провели, по-видимому, почти безвыходно в Африке, совершая иногда более или менее продолжительные вылазки лишь в некоторые соседствующие с ней районы. Одним из таких районов, несомненно, был Ближний Восток, а другим, более отдалённым — Кавказ, где недавно обнаружены хорошо сохранившиеся черепа и нижние челюсти ранних эректусов, а также их каменные орудия. Древность этих находок, происходящих с местонахождения Дманиси в Грузии, наверняка превышает миллион лет и, возможно, достигает даже 1,7 млн лет. Кроме того, возраст в 1,5 и даже в 2 млн лет предполагался некоторыми учёными ещё для индонезийских гомо эректус, но обоснованность этих датировок оставляет желать много лучшего, и большинство исследователей встречает сообщения такого рода скептически. Последний тому пример — дискуссия, развернувшаяся вокруг полученных в середине 90-х гг. датировок архантропов из яванских местонахождений Моджокерто и Сангиран, согласно которым их возраст составляет около 1,8–1,6 млн лет. Эти сенсационные результаты, казалось, требовали кардинального пересмотра устоявшихся представлений о месте и времени происхождения Homo erectus, а возможно, и рода гомо (Homo) в целом. Однако, как вскоре выяснилось, анализы, благодаря которым были получены столь древние даты, проводились по образцам пемзы, не связанным непосредственно с ископаемыми костями гоминид, а значит, любые выводы о возрасте последних в данном случае сомнительны. Скорее всего, эректус появился на Яве не ранее 1,2 млн лет назад. Таков же примерно возраст и древнейших достоверных следов пребывания гоминид в Китае. Ещё более поздний возраст имеют известные сейчас европейские гоминиды: самые ранние из них, если принимать в расчёт только хорошо проверенные данные, датируются концом нижнего плейстоцена, т. е., иначе говоря, временем около 800 тыс. лет назад. Впрочем, проникновение наших предков в Европу началось, несомненно, ещё в более древнее время, на что указывают находки в соответствующих геологических отложениях обработанных камней и других следов жизнедеятельности гоминид, но, судя по малочисленности таких материалов, первоначально это были не более чем короткие эпизодические визиты. Всерьёз же расширением ойкумены архантропы «занялись», по-видимому, не ранее конца нижнего плейстоцена, когда они уже довольно прочно обосновались в Восточной и Юго-Восточной Азии (рис. 19). Их путь в новые районы пролегал, скорее всего, через Ближний Восток, где существовал сухопутный мост, связывавший Африку с Евразией. Кроме того, не исключено, что в районах Гибралтара и Африканского Рога тоже были какие-то перемычки, делавшие возможными миграции из Северной Африки непосредственно в Европу, а из Восточной Африки на Аравийский полуостров и далее в глубинные районы Азии. 
Именно с Азией, кстати, связана история открытия вида эректусов, относящаяся ещё к концу позапрошлого века. Во многом она напоминает предшествовавшую ей историю открытия Генрихом Шлиманом Трои, только в роли немецко-российского коммерсанта Шлимана в данном случае выступил молодой голландский врач и анатом Евгений Дюбуа, а вдохновил его на поиски не Гомер, а уже упоминавшийся английский натуралист Альфред Рассел Уоллес. В отличие от Дарвина, полагавшего, что человек появился в Африке, Уоллес считал, что нашей прародиной была Восточная Азия и что останки гипотетического «недостающего звена» или, иначе говоря, обезьяночеловека могут быть с наибольшей вероятностью обнаружены именно там. Убеждённый аргументами английского учёного, Дюбуа поступил при первой возможности в качестве медика в армию и отправился на службу в Индонезию, бывшую тогда колонией Нидерландов. Поиски, предпринятые им в 80-е гг. на Суматре, успеха не принесли, но Дюбуа это не обескуражило, и он продолжил их на острове Ява. Здесь, на берегу р. Соло близ селения Триниль ему и посчастливилось сделать своё самое важное открытие. В октябре 1891 г. из земли была извлечена верхняя часть черепа, не похожая ни на что дотоле известное, а в следующем году появилась бедренная кость, напротив, очень близкая по форме к человеческой и явно принадлежавшая прямоходящему существу. Если первоначально Дюбуа приписал черепную кость обезьяне, то, соединив две находки, он понял, что нашёл именно то, что так долго искал. В 1894 г. он опубликовал работу, где на основании яванских материалов выделил род и вид питекантроп эректус или обезьяночеловек прямоходящий (Pithecanthropus erectus), использовав в первой части названия нового таксона имя, которое много раньше уже дал гипотетическому недостающему звену немецкий биолог Эрнст Геккель. И хотя к настоящему времени термин питекантроп (Pithecanthropus) почти вышел из употребления в научной литературе, поскольку те гоминиды, которых им обозначали, были перемещены в род Homo, в просторечии членов вида эректус, открытого Дюбуа, по-прежнему часто называют питекантропами. Несмотря на то что питекантропы существовали более миллиона лет, заселяли, по крайней мере, на поздних этапах своей истории, огромный ареал (всю Африку, юг Европы и большую часть Азии) и отдельные их популяции почти наверняка длительное время были разделены естественными барьерами, в анатомическом отношении они представляют собой сравнительно однородную группу. Конечно, по ряду признаков ископаемые материалы, относимые к виду эректус (Homo erectus), различаются между собой, но присущие тем или иным находкам специфические особенности не обнаруживают чёткой географической локализации, проявляясь в пространстве, скорее, чересполосно. Поэтому большинство антропологов не согласно с иногда высказываемым предложением оставить название Homo erectus только за восточноазиатскими архантропами, а африканских именовать иначе, и продолжают рассматривать первых и вторых как один вид. Новым весомым аргументом в пользу такой точки зрения стал череп возрастом около 1 млн лет, обнаруженный в 1997 г. в Среднем Аваше (Эфиопия), в отложениях геологической формации Бури. И по метрическим, и по качественным характеристикам эта восточноафриканская находка отличается от любого из черепов эректуса из Восточной Азии не больше, чем они сами различаются между собой.[27] Если отвлечься от ряда мелких деталей, то не будет преувеличением сказать, что для большинства представителей вида эректусов в общем характерен уже вполне человеческий скелет, который по многим параметрам, в том числе по размерам и пропорциям, очень близок к современному состоянию. Особенно показательно в этом смысле соотношение длины бедренной, плечевой и предплечевых костей. У шимпанзе оно равняется примерно 1:1, а у людей, хотя плечо имеет почти тот же абсолютный размер, бедро удлинено, а предплечье укорочено. Впервые эти человеческие пропорции фиксируются именно у эректусов около 1,5 млн лет назад (рис. 20). Вид эректусов отличается также от более ранних гоминид возросшими размерами тела и уменьшением различий по этому признаку между полами. Кроме того, уже у самых ранних эректусов (т. е. тех, кого причисляют ещё к виду эргастер (Homo ergaster), полностью отсутствуют какие бы то ни было черты, свидетельствующие о сохранении приспособленности к древесному образу жизни. В то же время ряд особенностей в строении таза, позвонков и костей конечностей ещё заметно отличает архантропов от более поздних гоминид. 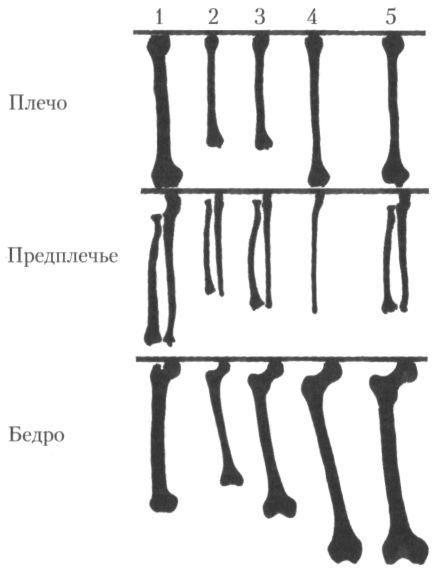
Череп на этой стадии эволюции человека изменяется не столь заметно, как остальной скелет, хотя тенденция к росту объёма мозговой полости, отчётливо наметившаяся уже у хабилисов, сохраняется. Это хорошо видно при сравнении, например, с черепом австралопитека африканского (рис. 21). Средний размер эндокрана у представителей вида эректус составляет около 1000 см3. Правда, сейчас ещё до конца не ясно, был ли процесс увеличения мозга постоянным и непрерывным, или же он носил скачкообразный характер. Об этом трудно судить с уверенностью не только потому, что черепов с хорошо сохранившимся мозговым отделом обнаружено пока не так много, но и потому, что большую их часть трудно точно датировать. В итоге одни исследователи считают, что на протяжении всей истории вида эректус происходило постепенное увеличение мозга гоминид, а другие убеждены, что скачок в объёме эндокрана имел место лишь на стадии ранних архантропов, а в последующий период вплоть до появления палеоантропов сколько-нибудь существенных изменений уже не было. 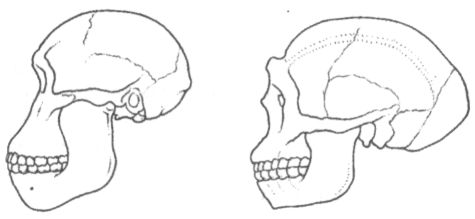
Несмотря на увеличение мозга, на черепе архантропов сохраняется много примитивных черт. Лобная кость у них низкая и покатая, с прямо выступающим надглазничным валиком, затылок угловатый, стенки черепа толстые, а наибольшая его ширина приходится не на среднюю часть, как у неандертальцев и современных людей, а на основание, как у австралопитеков. Для лицевого скелета характерно ещё заметное выступание вперёд ротовой части (хотя и не столь сильное, как у более ранних гоминид), а нижняя челюсть лишена подбородочного выступа и убегает назад. Жевательные зубы крупнее, чем у современных людей, резцы тоже, а у яванских эректусов иногда встречается даже диастема. В то же время, по относительному размеру зубов и нижней челюсти, а также по толщине эмали уже самые ранние архантропы отличаются от современных людей гораздо меньше, чем австралопитеки и хабилисы. Антропологические данные дают некоторые основания предполагать, что архантропы, по крайней мере, поздние, могли обладать зачатками речи. Во всяком случае, по размерам и строению челюстей и ротовой полости они гораздо ближе к современным людям, чем к обезьянам и, по-видимому, потенциально были способны совершать языком все движения, необходимые для того, чтобы успешно артикулировать гласные и согласные звуки. Это, конечно, ни в коем случае не доказывает, что речь, действительно, звучала на Земле уже на столь ранней стадии нашей эволюции, но в принципе такая возможность не исключена. Вместе с костями архантропов часто находят их каменные орудия. Иногда они очень похожи на орудия, которыми пользовались ещё хабилисы, а иногда сильно отличаются от них, превосходя их совершенством формы и обработки. Особенно эффектно выглядят крупные, симметричные, тщательно оббитые с двух сторон изделия с копьевидным или топоровидным концом, выполнявшие множество самых разных функций, но именуемые рубилами и колунами (рис. 22). Эти изделия типичны для ашеля или ашельской эпохи (термин происходит от названия местечка Сент-Ашель во Франции, где в позапрошлом веке было найдено много рубил), которая следует в археологической периодизации за олдувайской и охватывает период примерно от 1,5 млн до 200 тыс. лет назад. Правда, рубила и колуны находят почти исключительно только в западной части ареала расселения архантропов, тогда как в Восточной Азии они по непонятной пока причине встречаются крайне редко. В восточноазиатском регионе даже более поздние, чем эректус, гоминиды пользовались в основном орудиями, с технологической точки зрения близкими олдувайским. Причём этот факт до сих пор не получил сколько-нибудь удовлетворительного объяснения. Ещё более загадочным кажется то обстоятельство, что в Юго-Восточной Азии костные остатки эректусов вообще не сопровождаются какими бы то ни было искусственно изготовленными предметами. Возможно, они пока просто не найдены, хотя нельзя полностью исключить также, что первоначально гоминиды обходились в этом регионе без каменных орудий, найдя им более или менее равноценную замену в изделиях из иных, менее долговечных материалов, таких, например, как бамбук. 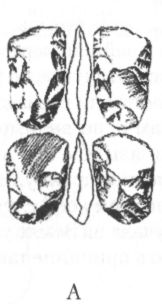 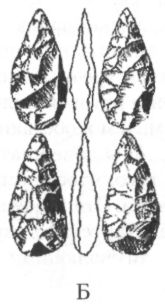
Эволюционная судьба различных географических популяций архантропов сложилась, вероятно, по-разному. Согласно преобладающей сейчас точке зрения, лишь африканские представители вида эректусов могли иметь прямое отношение к происхождению людей современного физического типа. Такие взгляды называют моноцентризмом, поскольку предполагается, что первоначальное становление гомо сапиенс было связано с одним, относительно ограниченным регионом (рис. 23А). В англоязычной литературе современную версию моноцентризма, утверждающую африканское происхождение всего современного человечества, называют также иногда теорией «Ноева ковчега». Противоположную позицию занимают исследователи, отстаивающие так называемую «мультирегиональную», или полицентристскую теорию, согласно которой эволюционное превращение эректусов в гомо сапиенс происходило повсеместно, т. е. и в Африке, и в Азии, и в Европе, при постоянном обмене генетическим материалом между популяциями, населявшими эти континенты (рис. 23Б). Хотя спор между моноцентристами и полицентристами, имеющий длительную историю, всё ещё далёк от завершения, инициатива сейчас явно в руках первых, а вторые занимают оборонительную позицию. Как мы увидим в следующей главе, накапливающиеся новые данные гораздо естественней укладываются в рамки теории, предполагающей, что неоантропы появились сначала в одном регионе (а именно в Африке), откуда распространились впоследствии по всей земле. Непосредственными же предшественниками их были палеоантропы — анатомически уже во многом не похожие на архантропов гоминиды второй половины среднего плейстоцена. 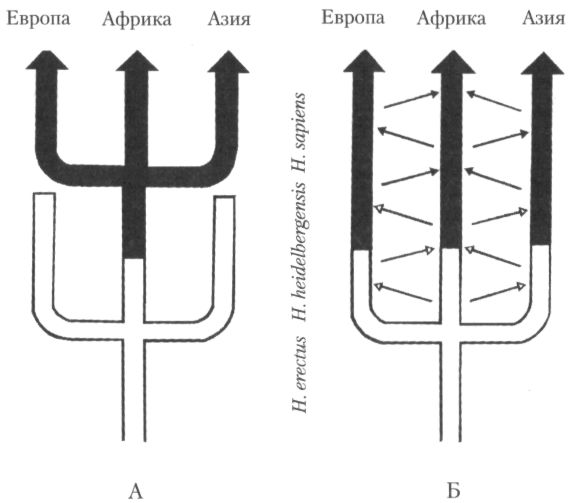
Палеоантропы Около полумиллиона лет назад или немного раньше в Африке и большинстве других регионов, где обитали эректусы, им на смену приходят гоминиды несколько иного типа. Являясь потомками архантропов и сохраняя ещё многие присущие им черты (низкий покатый лоб, развитые надбровные дуги, массивный подбородок без выступа и ряд других), они, тем не менее, повсеместно отличаются от своих предшественников общей формой черепа (рис. 24) и заметно возросшим объёмом мозговой полости. У этих гоминид, как правило, более широкий, чем у эректусов, лоб и более округлый затылок, лишённый свойственного архантропам угловатого выступа. Для них также характерно разрастание теменной области черепа. Размер эндокрана на этой стадии эволюции обычно превышает 1100 см3, а в отдельных случаях достигает даже размеров, типичных для современных людей (1400 см3 и более). 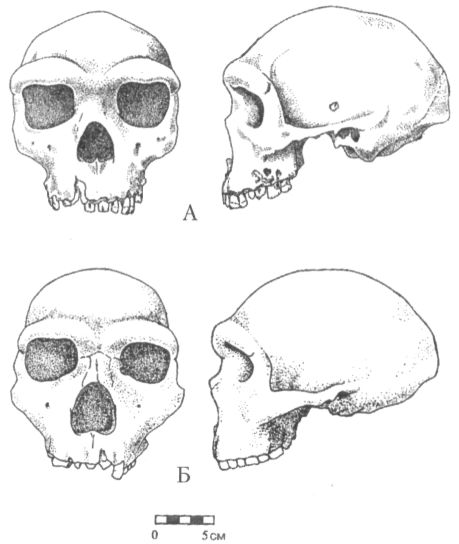
Хотя гоминиды, сменившие на эволюционной арене архантропов и господствовавшие на ней вплоть до появления людей современного физического типа, довольно сильно различались между собой и в пространстве, и во времени, условно их всё же было бы целесообразно объединять под общим именем, называя, например, палеоантропами. При этом речь должна идти, конечно, не о биологическом виде в собственном смысле, а о некоей стадии развития, эволюционной фазе, включающей множество форм, промежуточных по своему строению между эректусом и гомо сапиенс и не могущих быть отнесёнными ни к первому таксону, ни ко второму. Следует иметь в виду, что вплоть до недавнего времени эти гоминиды часто фигурировали в литературе как «архаичные» или «ранние» гомо сапиенс, что сильно запутывало терминологию и порой сбивало с толку даже некоторых исследователей, специализирующихся на изучении первобытности. Сейчас всех ранних палеоантропов относят обычно к виду человек гейдельбергский (как мы увидим далее, существуют и более дробные их классификации, придающие большее значение местным особенностям ископаемых материалов из разных регионов). Хотя этот вид был выделен почти сто лет назад, широкое признание он получил лишь в конце прошлого века. Тем самым были, наконец, достойно вознаграждены энтузиазм и прозорливость его первооткрывателя, немецкого антрополога Отто Шетензака, который, подобно Дюбуа, в течение долгих лет целенаправленно искал кости ископаемого человека. Правда, в отличие от Дюбуа, Шетензаку не пришлось отправляться за находкой его жизни на край света: он сделал своё открытие неподалёку от Гейдельберга (Германия), где преподавал в университете. В нескольких километрах от этого города, близ деревни Мауэр, располагался песчаный карьер, который ещё с середины XIX в. был известен как место многочисленных находок костей давно вымерших плейстоценовых млекопитающих — древнего слона, этрусского носорога, разных видов медведей, лошадей, и т. д. Шетензак на протяжении 20 лет регулярно наведывался в Мауэр в надежде, что в один прекрасный день среди палеонтологических находок, извлекаемых из земли рабочими карьера, могут обнаружиться и останки очень древней формы человека. В октябре 1907 г. такой день, действительно, наступил. На глубине 24 м от поверхности одним из рабочих была найдена, наряду с костями упомянутых выше животных, массивная нижняя челюсть с хорошо сохранившимися зубами, которая явно принадлежала гоминиду (рис. 25). Спустя год Шетензак опубликовал книгу об этой находке, где и был провозглашён новый вид — человек гейдельбергский (Homo heidelbergensis). Ещё через пять лет первооткрыватель гейдельбергского человека умер. 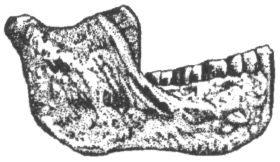
По многим чертам своего строения челюсть из Мауэра выглядит очень примитивной, и долгое время её приписывали питекантропу. Однако плавные параболические очертания зубной дуги и, главное, размеры и форма самих зубов сближают её обладателя с гомо сапиенс и другими поздними гоминидами. Поэтому, когда во второй половине прошлого века в Африке, Европе и Азии были обнаружены в довольно большом количестве новые скелетные останки, промежуточные по своей анатомии между видом эректус, с одной стороны, и современными людьми и неандертальцами, с другой, вполне естественным оказалось их зачисление в одну группу с гейдельбергским человеком. Эту группу стали теперь рассматривать как отдельный вид, не смешивая его более с питекантропами и вернув ему название Homo heidelbergensis, предложенное Шетензаком. Кроме Мауэра, наглядные свидетельства этой стадии эволюции человека, в виде ископаемых черепов и других частей скелета, найдены на африканских местонахождениях Омо, Бодо (оба в Эфиопии), Ндуту, Нгалоба, Эяси (все в Танзании), Брокен-Хилл (Замбия) и ряде других, а также на Ближнем Востоке — Зуттиех в Израиле, в Южной Азии — Нармада в Индии, Восточной Азии, Дали, Мапа, Сюцзияо в Китае и др., в Европе — Петралона в Греции, Араго во Франции и т. д. Возраст большинства перечисленных находок не определён сколько-нибудь точно, но в целом они могут быть размещены в хронологическом диапазоне примерно от 600 до 150 тыс. лет назад. В культурном отношении палеоантропы, за исключением их самых поздних разновидностей, очень мало отличались от архантропов. Они пользовались такими же орудиями, жили целиком за счёт собирательства и охоты и часто передвигались с места на место в поисках пищи. Единственная сфера поведения, где, возможно, произошли очень важные изменения по сравнению с предшественниками — это общение, коммуникация. Судя по некоторым анатомическим признакам, многие палеоантропы уже вполне могли владеть членораздельной речью. Как известно, строение основания черепа (базикраниума) коррелирует с определёнными особенностями строения мягких тканей верхних дыхательных путей. Исследованиями на ряде млекопитающих было показано, что существует связь между степенью изогнутости основания черепа и положением гортани в горле. При слабо изогнутом базикраниуме гортань расположена высоко, при сильно изогнутом — низко. Последняя черта характерна для людей, причём у детей до двух лет гортань расположена также высоко как у животных (что даёт им и животным возможность есть и дышать практически одновременно), и лишь в 3 года начинает опускаться (что позволяет лучше и разнообразнее артикулировать звуки, но создаёт риск подавиться). С целью реконструировать положение гортани изучались базикраниумы ископаемых гоминид. Было установлено, что австралопитеки в этом отношении гораздо ближе к человекообразным обезьянам, чем к современным людям. Изменения в современном направлении начались у ранних эректусов: анализ черепа KNM-ER 3733 возрастом около 1,5 млн лет выявил зачаточный изгиб базикраниума. А вот у ранних палеоантропов (черепа из Петралоны и Брокен-Хилл) фиксируется уже полный изгиб, близкий к характерному для черепов современных людей, что означает и такое же низкое положение гортани в горле. Трудно представить, что риск подавиться, связанный с такого рода анатомическими изменениями, был их единственным результатом и не компенсировался с самого начала иной, полезной функцией (или функциями). Поэтому резонно предполагать, что те гоминиды, у которых гортань уже была расположена достаточно низко, не просто имели возможность членораздельной речи, но и пользовались этой возможностью. Можно отметить также, что палеоантропы наверняка систематически пользовались огнём. Умели ли они его добывать и, если да, то каким способом, пока неизвестно, но так или иначе, а в среднем плейстоцене эта природная сила уже служила человеку. На стоянках Вертешселлеш (Венгрия), Бильцингслебен (Германия), Терра-Амата (Франция) и ряде других найдены следы костров, горевших примерно 300–400 тыс. лет назад. В несколько более поздний период очаги и кострища на поселениях палеоантропов становятся вполне обычным явлением, распространённым от Южной Африки до Европы. Хотя, как уже было сказано, по мнению большинства антропологов, вид человека гейдельбергского включает в себя всех ранних палеоантропов, независимо от того, найдены ли они в Южной Африке, в Восточной Азии, или в Западной Европе, эти гоминиды далеко не одинаковы в анатомическом отношении. Они могут быть разделены на несколько географических, а также и хронологических групп, которым некоторые исследователи склонны даже придавать статус самостоятельных видов. Действительно, вполне естественно, что через какое-то время после широкого расселения эректусов за пределы Африки между разбросанными в пространстве человеческими популяциями стало постепенно накапливаться всё больше и больше различий. Непохожие природные условия Африки, Восточной Азии и Средиземноморья предъявляли к обитавшим в этих регионах гоминидам разные требования, отбор работал в разных направлениях и с разной интенсивностью. Как следствие этого, эволюционные изменения должны были вести к формированию специфических особенностей в поведении и анатомии каждого из географических вариантов если не архантропов, то, по крайней мере, сменивших их палеоантропов. Конечно, вполне возможно, что процесс расхождения в какой-то мере сглаживался за счёт обмена генами, которые даже при скрещивании только между соседними популяциями со временем могли распространяться на многие тысячи и десятки тысяч километров, но вряд ли этого было достаточно для поддержания на протяжении всего среднего плейстоцена биологического единства гоминид, живших на разных континентах. Даже если допустить, что они, как полагают отдельные антропологи, оставались членами одного вида в репродуктивном смысле, т. е. сохраняли способность скрещиваться и давать при этом нестерильное потомство, углубляющиеся различия в анатомии и в поведении, а также географическая изоляция вели, вероятно, к тому, что на практике эта способность реализовывалась нечасто. Подобная ситуация характерна для многих современных животных, например, для волка и собаки. Ещё один пример — африканские львы и бенгальские тигры. Внешне столь непохожие и относимые к разным видам рода пантера (Panthera), они, тем не менее, не разделены ещё бесповоротно барьером репродуктивной изоляции, что успешно доказывают в зоопарках. Палеоантропологические материалы свидетельствуют о том, что в Европе, начиная уже с рубежа нижнего и среднего плейстоцена, местные гоминиды приобретают всё больше признаков, отличающих их от африканских сородичей и современников. Правда, бесспорных останков архантропов на нашем континенте пока практически нет. Единственное возможное исключение представляет собой неполная черепная коробка, обнаруженная в 1994 г. в городке Чепрано неподалёку от Рима в отложениях возрастом порядка 800 тыс. лет. Её сначала отнесли, хотя и не без оговорок, к виду эректусов, а впоследствии предложили рассматривать как переходную форму между видом эректус и человеком гейдельбергским. Костные останки древнейших известных нам европейских палеоантропов настолько своеобразны, что иногда их даже отделяют от человека гейдельбергского и включают в особый вид, получивший название антецессор (Homo antecessor). Этот вид, по мнению настаивающих на его независимом статусе испанских антропологов, представлен сейчас останками как минимум шести гоминид. Все они были найдены в пещере Гран Долина в Атапуэрке на северо-востоке Испании и имеют древность около 800 тыс. лет, относясь, таким образом, к самому концу нижнего плейстоцена. Остатки гоминид из Гран Долины происходят из одного и того же геологического слоя. Они залегали в нём вперемешку с костями животных и каменными изделиями. Интересно, что при этом большинство человеческих костей несёт на своих поверхностях точно такие же следы надрезов и ударов орудиями, какие имеются на прочих фаунистических остатках. Не исключено, что в данном случае перед нами свидетельства существования каннибализма. Коллекция костей довольно представительна: она включает несколько десятков зубов, обломок нижней челюсти, хорошо сохранившуюся лицевую часть черепа, позвонки, рёбра, ключицы и кости конечностей. По многим важным признакам (пропорции предкоренных зубов, большая ширина лобной кости, современная морфология лицевого скелета и др.) эти древнейшие европейцы сильно отличались от живших в одно время с ними в Африке эректусов, приближаясь к гораздо более поздним гоминидам. Те исследователи, которые признают антецессора в качестве самостоятельного вида, часто рассматривают его как связующее звеном между эргастером и человеком гейдельбергским, отводя при этом поздним архантропам (эректусам) роль тупиковой ветви в нашей эволюции. Предполагается также, что антецессор мог быть последним общим предком неандертальцев и современных людей. Согласно другой точке зрения, гоминиды из Гран Долины относятся к виду человека гейдельбергского, стоят у истоков линии, ведущей к неандертальцам, и не имеют прямого отношения к происхождению гомо сапиенс. Картина, конечно, в итоге получается крайне запутанная, и пройдёт, вероятно, ещё немало времени, прежде чем она будет прояснена. Среди европейских палеоантропов, следующих по времени за гоминидами из Гран Долины и Чепрано, достаточно отчётливо выделяются две группы. Это так называемые пренеандертальцы, которых почти все исследователи относят теперь к виду человека гейдельбергского, и произошедшие от них «классические» неандертальцы, выделяемые в самостоятельный вид неандерталец (Homo neanderthalensis). Пренеандертальцы жили в среднем плейстоцене и заселяли весь юг и запад Европы от Балкан до Британских островов и от Рейна и Дуная до Пиренейского полуострова. Помимо уже упоминавшейся челюсти из Мауэра, их останки включают черепа из Петралоны (Греция), Араго (Франция), Штейнгейма (Германия) и множество других находок. Сменившие пренеандертальцев классические неандертальцы появляются приблизительно на рубеже среднего и позднего плейстоцена, т. е. 130–150 тыс. лет назад. Некоторые исследователи, правда, предполагают, что в качестве наиболее древних представителей вида неандертальцев следует, возможно, рассматривать уже гоминид из Сванскомба в Англии и Сима де лос Хэсос в Испании, живших 300–400 тыс. лет назад. Местом происхождения неандертальцев наверняка была Европа, но впоследствии они значительно расширили ареал своего обитания, заселив сначала Западную Азию, а затем и более отдалённые районы на востоке вплоть до южной Сибири (рис. 26). Этот вид существовал на протяжении как минимум 100 тысячелетий: последние его представители жили в ряде районов Евразии ещё 30 тыс. лет назад и даже несколько позднее. 
Термин «неандерталец» происходит от названия долины Неандер или Неандерталь («таль» по-немецки «долина») в Германии, близ Дюссельдорфа. Здесь находится пещера Фельдгофер, где ещё в 1856 г. были обнаружены многочисленные кости одного из представителей этого вида гоминид, а именно: черепная крышка, конечности, фрагменты рёбер, таза и т. д. Правда, как выяснилось позже, останки неандерталецев неоднократно находили ещё в первой половине XIX в., сначала в Бельгии, а потом в Испании (точнее, на Гибралтаре, принадлежавшем и всё ещё принадлежащем Англии), но тогда их не смогли оценить должным образом. Находки же из Фельдгофер, опубликованные немецким анатомом Германом Шафгаузеном в 1857 г., т. е. всего за два года до выхода в свет «Происхождения видов» Дарвина, сразу же привлекли внимание учёного мира, и шесть лет спустя, в 1863 г., английский геолог Вильям Кинг предложил для древнего обитателя Неандерталя название Homo neanderthalensis. Это название довольно символично, поскольку слово «неандер» означает буквально «новый человек». Оно представляет собой греческую версию имени жившего в Дюссельдорфе в XVII в. композитора и священнослужителя Йоахима Ноймана. Относительно костей из Неандерталя долгое время велись жаркие споры. Многие авторитетные учёные XIX столетия не признавали их в качестве останков человека другого вида, высказывая при этом порой идеи, которые сейчас трудно воспринимать без улыбки. Предполагалось, например, что наличие массивных надбровных дуг на черепе из пещеры Фельдгофер (рис. 27), столь нехарактерное для современных людей, объясняется его принадлежностью русскому казаку, участвовавшему в боях с наполеоновской армией в Европе и постоянно хмурившемуся от боли, которую причиняли ему полученные в сражениях раны. Некоторые авторы решали проблему ещё радикальней, утверждая, что череп просто принадлежал идиоту с патологическими отклонениями в строении скелета. Впрочем, уже к началу прошлого столетия, благодаря ряду новых находок, похожих на кости, описанные Шлафгаузеном, но при этом сопровождавшихся ископаемой фауной и каменными орудиями, сомнения относительно древнего возраста неандертальца были рассеяны. Кстати, совсем недавно, два года назад, останки ещё двух представителей неандертальского вида человека были найдены и в самой долине Неандерталь. Кости рук принадлежали, вероятно, женщине, а молочные зубы ребёнку 10–12 лет. Удалось определить также, что эти люди жили примерно за 40 тыс. лет до наших дней.[28] 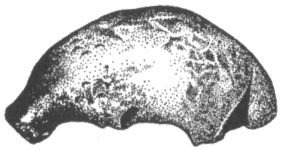
Сейчас неандертальцы являются, пожалуй, наиболее полно изученной группой гоминид, если не считать людей современного физического типа. Разумеется, мы знаем о них далеко не всё, что хотелось бы, но всё же намного больше, чем, скажем, об австралопитеках, или даже о ранних палеоантропах. Их останки, представленные в одних случаях разрозненными костями, а в других полными или почти полными скелетами, были найдены на многих археологических памятниках Европы и западной Азии. Особенно часто они встречаются на пещерных стоянках эпохи среднего палеолита, таких, как Ля Феррасси и Ля Кина во Франции, Спи в Бельгии, Заскальная в Крыму, Мезмайская на Северном Кавказе, Кебара, Амуд и Табун в Израиле, Шанидар в Ираке, Тешик-Таш в Узбекистане и др. В ряде случаев хорошая сохранность костяков объясняется тем, что они были захоронены в специально вырытых ямах или естественных углублениях, представляющих собой, таким образом, древнейшие достоверно известные погребения. При относительно невысоком среднем росте (165–170 см) для неандертальцев характерна массивность костей скелета, свидетельствующая о большой мышечной массе. Они, несомненно, обладали значительной физической силой, но красавцами их, если руководствоваться сегодняшними эстетическими мерками, назвать было бы трудно. Впрочем, иной неандерталец, одетый в современный костюм, побритый и подстриженный (лучше — наголо), пожалуй, вполне органично смотрелся бы где-нибудь на Невском проспекте или на Бродвее в Нью-Йорке, и его появление там не вызвало бы особого ажиотажа (рис. 28). Свой низкий и покатый лоб он мог бы спрятать под шляпой, крупные широко расставленные орбиты глаз с развитыми надбровными дугами скрыть за тёмными очками, ну а нижнюю челюсть без подбородочного выступа просто выпятить вперёд, выставив напоказ — на Невском и не такое увидишь! Коренные зубы у неандертальцев были меньше, чем у других гоминид, за исключением H. sapiens, а резцы, наоборот, крупнее, чем у предшественников и гораздо крупнее, чем у людей современного физического типа. Черепная коробка у них длинная и широкая, причём наибольшая ширина приходится на среднетеменную часть. Специфическими чертами анатомии черепа, отличающими неандертальцев от всех современных им и более поздних гоминид, считаются его субсфероидная форма (если смотреть сзади), заметное выступание вперёд лицевого скелета, наличие затылочного валика и жевательного гребня за слуховым отверстием (рис. 29). Ещё одним признаком, имеющимся у почти всех неандертальцев и отсутствующим у большинства представителей других видов рода Homo, является так называемый «ретромолярный пробел» — пустое пространство между задними коренными зубами и восходящей ветвью нижней челюсти. 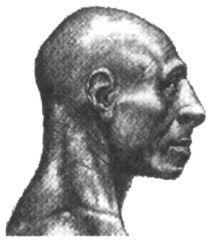
 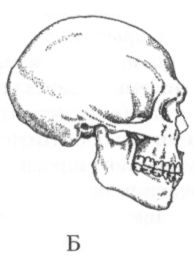
Несмотря на общий «архаизм» облика неандертальцев, абсолютная величина мозговой полости у них и в среднем (1400–1500 см3), и по размаху крайних значений практически не отличалась от таковой у современных людей, даже несколько превосходя её. Что же до величины относительной, то здесь некоторая — пусть и незначительная — разница в нашу пользу всё же, видимо, существовала, хотя достигнута она была, скорее всего, не за счёт увеличения самого мозга, а за счёт уменьшения массы тела. Впрочем, абсолютный размер мозга гомо сапиенс также слегка изменялся от эпохи к эпохе, что, по всей вероятности, в значительной степени было связано с эпохальными изменениями размеров тела. В частности, показано, что в Европе, начиная с конца ледникового периода, люди постепенно становились всё меньше, и абсолютный размер мозга в этот период также уменьшался. Не исключено, что этот процесс продолжался непрерывно в течение очень долгого времени. По мнению некоторых антропологов, основанному на изучении тысяч человеческих черепов из разных регионов, уменьшение размера мозга шло безостановочно на протяжении всего голоцена и минимального для гомо сапиенс значения этот показатель достиг как раз в историческое время. Очень сложен вопрос об окончательной судьбе неандертальцев, тесно связанный с проблемой происхождения гомо сапиенс. Уже столетие он находится в центре дискуссий учёных, занимающихся поисками генеалогических корней современного человечества. Ответы на него предлагались самые разные, часто взаимоисключающие. С одной стороны, долгое время казалось очевидным, что во времени неандертальцы являются непосредственными предшественниками гомо сапиенс и, таким образом, их наиболее вероятными предками. С другой стороны, многие антропологи отмечали, что по целому ряду черт своего анатомического строения обе эти разновидности гоминид довольно-таки резко различаются, а хронологический промежуток, разделяющий их, слишком мал, чтобы можно было предполагать плавное эволюционное превращение одной формы в другую. В конце концов выяснилось, что на самом деле никакого промежутка вообще не было, и, скорее всего, неандертальцы и люди современного анатомического типа появились примерно одновременно, хотя и в разных частях света. Это означает, что неандертальцы не были нашими прямыми предками и, если и внесли какой-то вклад в генофонд нынешнего человечества в результате ассимиляции, то весьма скромный. Правда, некоторые исследователи считают, что, по крайней мере, в Европе вид неандертальцев постепенно эволюционировал в вид гомо сапиенс, но убедительно показать наличие преемственности анатомических признаков между первыми европейскими неоантропами и неандертальцами пока никому не удалось, и более вероятным кажется отсутствие таковой. Против подобной преемственности свидетельствует и твёрдо установленный факт довольно длительного сосуществования этих двух форм гоминид в Западной Европе, где они жили бок о бок на протяжении как минимум пяти, а то и десяти тысяч лет, а также результаты уже упоминавшихся палеогенетических исследований. К настоящему времени удалось извлечь фрагменты ДНК из костей четырёх неандертальцев, в том числе несостоявшегося «казака» из долины Неандер и грудного младенца из пещеры Мезмайская на Северном Кавказе. Их сопоставление с ДНК современных людей показало, что генетическая «дистанция» между неандертальцами и гомо сапиенс в три с лишним раза превышает разницу между ныне существующими расовыми группами человечества (негроидами, монголоидами и европеоидами). Кроме того, выяснилось, что ДНК неандертальцев ничуть не ближе по своему строению к ДНК европейцев, чем к ДНК коренного населения остальных частей света, а между тем, если допустить, что гипотеза о преемственности между двумя видами в Европе справедлива, то следовало бы ожидать совсем иную картину. Всё сказанное заставляет считать, что гомо сапиенс пришли в Европу извне, а её исконные обитатели, неандертальцы, скорее всего, не выдержали конкуренции с пришельцами и разделили незавидную участь многих других видов млекопитающих, исчезнувших в конце плейстоцена с лица земли (мамонт, шерстистый носорог и т. д.). Несколько подробней о том, почему это могло произойти, ещё будет говориться в следующей главе. К началу верхнего плейстоцена, т. е. как раз к тому времени, когда в Европе появляются неандертальцы, в Африке заканчивается в основном сложение комплекса анатомических признаков, характеризующих людей современного физического типа. Предшественниками последних на африканском континенте были местные палеоантропы, которых обычно относят вместе с их европейскими современниками к виду человека гейдельбергского, но иногда выделяют в особые виды гомо родезиенс, или человек родезийский (Homo rhodesiensis) и гомо гельмей (Homo helmei). В первый из них при этом включают ранних африканских палеоантропов, а во второй поздних. О них также будет ещё сказано несколько слов в главе 5, при рассмотрении проблемы происхождения гомо сапиенс. Ещё один (как минимум) вариант человечества мог формироваться в Восточной и Юго-Восточной Азии. Весьма специфический характер культурных адаптаций в этом регионе на протяжении почти всего палеолита позволяет предполагать, что и биологическая эволюция здесь должна была идти несколько иными путями, нежели в Африке и Европе. Однако ввиду скудости и недостаточной изученности антропологических материалов, точно определить направление развития и степень обособленности восточноазиатских палеоантропов пока невозможно. Стоит, однако, отметить, одно очень интересное обстоятельство, лишний раз показывающее, насколько по-разному могла складываться история представителей одного и того же вида гоминид в разных частях света. Судя по датировкам, полученным недавно двумя разными методами, для тех слоёв местонахождений Нгандонг и Самбунгмакан (Ява), где были найдены останки вида эректус, этот вид мог сосуществовать здесь какое-то время с гомо сапиенс. На основе фаунистических и геоморфологических данных давно уже высказывалось предположение, что архантропы на Яве дожили до позднего плейстоцена. Теперь об этом свидетельствуют и абсолютные даты, варьирующие от 25 до 55 тыс. лет назад. Напрашивается аналогия с Европой, где гомо сапиенс также сосуществовали в течение какого-то времени с гоминидами иного типа, хотя и гораздо более «продвинутого» по сравнению с видом эректус. Примечания:[2] По-гречески «нос» — ???, слово «?????» — форма этого существительного в родительном падеже. (Примеч. ред.) [22] Этот термин уже используют некоторые авторы, например: Brumann С. On culture and symbols // Current Anthropology. 2002. Vol. 43. № 3. P. 510. Кроме того, применительно к шимпанзе, родовое название которых — Pan, стали говорить также о культурной пантропологии (Whiten et al. Cultural panthropology // Evolutionary Anthropology. 2003. Vol. 12. № 2. P. 92–105). [23] Майр Э. Зоологический вид и эволюция. М., 1968. С. 502. [24] Tobias P. V. Biological equipment, environment and survival in the Australopithecine world // Hominid Evolution. Lifestyles and Survival Strategies. Gelsenkirchen, 1999. P. 55–71. [25] Tobias P. V. The brain of the first hominids // Origins of the Human Brain. Oxford, 1995. P. 77. [26] Krings М. et al. A view of Neandertal genetic diversity // Nature Genetics. 2000. Vol. 26. P. 144–146. [27] Asfaw В. et al. Remains of Homo erectus from Bouri, Middle Awash, Ethiopia // Nature. 2002. Vol. 416. P. 317–320. [28] Schmitz R. W. et al. The Neandertal type site revisited: Interdisciplinary investigations of skeletal remains from the Neander Valley, Germany // Proceedings of the National Academy of Sciences USA. 2002. Vol. 99. № 20. P. 13342–13347. |
|
||
|
Главная | Контакты | Нашёл ошибку | Прислать материал | Добавить в избранное |
||||
|
|
||||