|
||||
|
|
Глава 4. Морфогенетические поля 4.1. Морфогенетические зародыши Морфогенез не происходит в вакууме. Он может начаться только с уже организованной системы, которая служит морфогенетическим зародышем. В процессе морфогенеза новая морфическая единица более высокого уровня возникает вокруг этого зародыша под влиянием специфического морфогенетического поля. Так как же это поле соединяется с морфогенетическим зародышем, чтобы началось развитие? Ответ может состоять в том, что как соединение материальных систем с гравитационными полями зависит от их массы, а с электромагнитными полями — от их электрического заряда, так и соединение систем с морфогенетическими полями зависит от их формы. Таким образом, морфогенетический зародыш окружается морфогенетическим полем благодаря тому, что он имеет свою характерную форму. 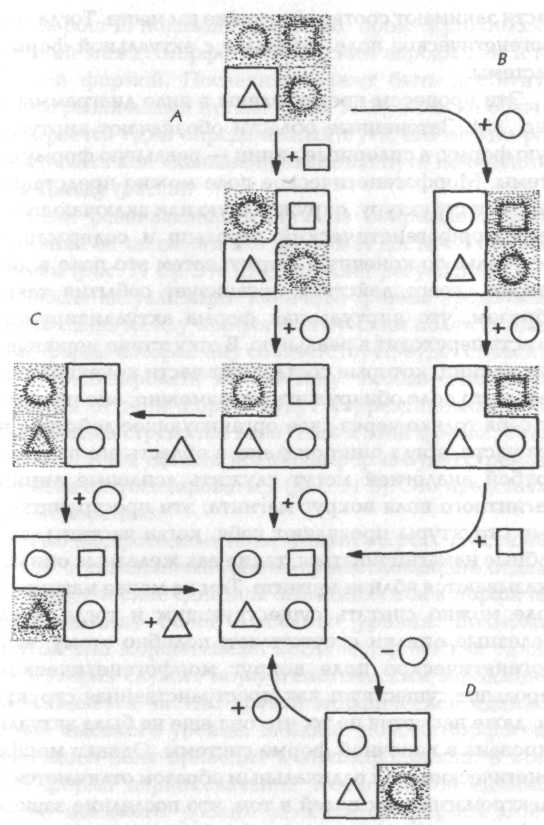 Рис. 11. Диаграмма, представляющая развитие системы из морфогенетического зародыша (треугольник) по нормальной хреоде (А). Показаны также альтернативный морфогенетический путь развития (В), регуляция (С) и регенерация (D). Затененные области представляют виртуальные формы внутри морфогенетического поля Морфогенетический зародыш является частью системы, которая должна возникнуть. Поэтому часть морфогенетического поля системы соответствует этому зародышу. Однако остальная часть поля еще не занята или не заполнена; она содержит виртуальную форму законченной системы, которая становится реальной (актуальной) только тогда, когда все ее материальные части занимают соответствующие им места. Тогда морфогенетическое поле совпадает с актуальной формой системы. Эти процессы представлены в виде диаграммы на рис. 11. Затененные области обозначают виртуальную форму, а сплошные линии — реальную форму системы. Морфогенетическое поле можно представить себе как структуру, окружающую или включающую в себя морфогенетический зародыш и содержащую виртуальную конечную форму; затем это поле в диапазоне своего действия организует события таким образом, что виртуальная форма актуализируется, то есть переходит в реальную. В отсутствие морфических единиц, которые составляют части конечной системы, это поле обнаружить невозможно; оно проявляет себя только через свое организующее действие на эти части, когда они попадают в область его влияния. Грубой аналогией могут служить «силовые линии» магнитного поля вокруг магнита; эти пространственные структуры проявляют себя, когда частицы, способные намагничиваться, такие как железные опилки, оказываются вблизи магнита. Тем не менее магнитное поле можно считать существующим и тогда, когда железные опилки отсутствуют; подобно этому, морфогенетическое поле вокруг морфогенетического зародыша существует как пространственная структура, даже несмотря на то, что она еще не была актуализирована в конечной форме системы. Однако морфогенетические поля радикальным образом отличаются от электромагнитных полей в том, что последние зависят от фактического (актуального) состояния системы — от распределения и движения заряженных частиц, — тогда как морфогенетические поля соответствуют потенциальному состоянию развивающейся системы и уже присутствуют раньше, чем система принимает свою конечную форму.[105] На рис. 11 показано несколько промежуточных состояний между морфогенетическим зародышем и конечной формой. Последняя может быть достигнута также различными путями {рис. 11 В), но если обычно выбирается один определенный путь, его можно рассматривать как «канализированный путь изменения», или хреоду (рис. 5). Если развивающаяся система повреждена путем удаления ее части, она все же может достичь конечной формы (рис. 11 С). Это представляет регуляцию. После актуализации конечной формы продолжающаяся связь между морфогенетическим полем и системой, форма которой ему соответствует, будет стремиться стабилизировать эту систему. Любые отклонения системы от этой формы будут корректироваться, так как система стремится вернуться к этой форме. А если часть системы удалить, конечная форма будет стремиться снова актуализироваться (рис. 11 D). Это представляет регенерацию. Тип морфогенеза, показанный на рис. 11, существенно агрегативный: ранее существовавшие отдельно морфические единицы объединяются в морфическую единицу более высокого уровня. Возможен другой тип морфогенеза, когда морфическая единица, которая служит морфогенетическим зародышем, уже является частью другой морфической единицы более высокого уровня. Влияние нового морфогенетического поля приводит к трансформации, в которой форма первоначальной морфической единицы более высокого уровня заменяется формой новой морфической единицы. Большинство видов химического морфогенеза относится к агрегативному типу, тогда как биологический морфогенез предполагает обычно комбинацию трансформативного и агрегативного процессов. Примеры мы рассматриваем в следующих разделах. 4.2. Химический морфогенез Агрегативные морфогенезы с нарастающей интенсивностью осуществляются в неорганических системах при понижении температуры: когда плазма охлаждается, субатомные частицы агрегируют в атомы; при более низкой температуре атомы агрегируют в молекулы; затем молекулы конденсируются в жидкости; и наконец, жидкости кристаллизуются. В состоянии плазмы обнаженные атомные ядра можно рассматривать как морфогенетические зародыши атомов; они связаны с атомными морфогенетическими полями, которые содержат виртуальные орбитали электронов. В одном смысле эти орбитали не существуют, но в другом — они обладают реальностью, которая выявляется в остывающей плазме, когда они актуализируются путем захвата электронов. Электроны, которые были захвачены атомными орбиталями, могут быть смещены оттуда воздействием внешней энергии или путем введения виртуальной орбитали с более низкой потенциальной энергией. В последнем случае они теряют дискретный квант энергии, который излучается как фотон. В атомах со многими электронами каждая орбиталь может содержать только два электрона (с противоположными спинами); таким образом, в остывающей плазме электронами заполняются сначала виртуальные орбитали с наиболее низкой энергией, затем орбитали с ближайшими к ним низшими энергиями и так далее, до тех пор пока не реализуется (актуализируется) полная атомная форма вокруг морфогенетического зародыша ядра. Атомы, в свою очередь, являются морфогенетическими зародышами молекул, а малые молекулы — зародышами больших молекул. Химические реакции включают либо агрегацию атомов и молекул в большие молекулы — например, образование полимеров, — либо расщепление больших молекул на меньшие или на атомы и ионы, которые затем могут агрегировать с другими атомами или ионами, например в процессе горения: под действием внешней энергии молекулы распадаются на атомы и ионы, которые затем соединяются с кислородом с образованием малых, простых молекул, таких как Н2О и СО2. Эти химические изменения включают актуализацию виртуальных форм, связанных с атомами или молекулами, которые действуют как морфогенетические зародыши. Мысль о том, что молекулы имеют виртуальные формы ранее, чем они актуализируются, особенно ясно иллюстрируется тем хорошо известным фактом, что совершенно новые соединения сначала могут быть «спроектированы» на основе эмпирически определенных принципов химического взаимодействия, а затем уже фактически синтезированы химиками-органиками. В этих лабораторных синтезах одна стадия следует за другой; на каждой стадии определенная молекулярная форма служит морфогенетическим зародышем для следующей виртуальной формы, которая должна быть синтезирована, и в конечном итоге образуется форма совершенно новой молекулы. Если представление о химических реакциях как о морфогенетических процессах кажется несколько искусственным, то следует вспомнить о том, что действие катализаторов, как неорганических, так и органических, в значительной степени зависит от их формы. Например, ферменты, специфические катализаторы множества биохимических реакций, имеют поверхности, канавки, выемки или впадины, которым соответствуют реагирующие молекулы со специфичностью, часто сравниваемой с тем, как ключ соответствует замку. Каталитический эффект ферментов в большой степени зависит от того способа, которым они удерживают реагирующие молекулы в положениях, нужных для того, чтобы реакция произошла. (В свободном растворе случайные столкновения молекул происходят во всех возможных ориентациях, большинство из которых непригодны для реакции.) Детали химического морфогенеза неясны отчасти из-за их большой скорости, отчасти потому, что промежуточные формы могут быть весьма нестабильны, а также потому, что конечные изменения состоят из вероятностных квантовых скачков электронов между орбиталями, которые образуют химические связи. Виртуальная форма молекулы, которая должна возникнуть, запечатлена в морфогенетическом поле, связанном с атомным или молекулярным морфогенетическим зародышем; когда другой атом или молекула приближаются в подходящей ориентации, форма образующейся молекулы актуализируется посредством квантовых скачков электронов на орбитали, которые ранее существовали лишь как виртуальные формы; в то же время при этом освобождается энергия, обычно в виде теплового движения. Роль морфогенетического поля в этом процессе является, так сказать, энергетически пассивной, но морфологически активной; оно создает виртуальные структуры, которые затем актуализируются, по мере того как морфические единицы низшего уровня «защелкиваются» («slot» or «snap») в эти структуры, освобождая при этом энергию. Любой атом или молекула данного типа могут участвовать во многих видах химических реакций и поэтому являются потенциальными зародышами множества различных морфогенетических полей. Эти поля можно представить как возможности, «парящие» вокруг них. Однако атом или молекула могут не выполнить свою роль зародыша определенного морфогенетического поля до тех пор, пока к нему или к ней не приблизится подходящий атом или молекула, возможно, благодаря их специфическим электромагнитным или иным воздействиям. Морфогенез кристаллов отличается от такового атомов и молекул тем, что определенный рисунок атомной или молекулярной организации повторяется неограниченно. Морфогенетический зародыш создается самим этим рисунком. Хорошо известно, что введение «семян» или «ядер» кристалла соответствующего типа значительно ускоряет кристаллизацию переохлажденных жидкостей или перенасыщенных растворов. В отсутствие таких семян или ядер морфогенетические зародыши кристаллов появляются только тогда, когда атомы или молекулы случайно занимают подходящие положения относительно друг друга вследствие теплового движения. Когда зародыш присутствует, виртуальные формы копий структуры решетки, задаваемой этим морфогенетическим полем, распространяются в разные стороны от поверхностей растущего кристалла. Подходящие свободные атомы или молекулы, которые приближаются к этим поверхностям, захватываются ими и «защелкиваются» в нужное положение; опять-таки при этом выделяется тепловая энергия. Осеменение, или нуклеация, переохлажденных жидкостей может также проводиться, хотя и менее эффективно, путем введения малых фрагментов чужеродных веществ: например, химики часто скребут по краям пробирок, чтобы засеять растворы кусочками стекла. Эти кусочки служат поверхностями, позволяющими легче находить наиболее подходящие относительные положения тем атомам или молекулам, которые образуют истинный морфогенетический зародыш кристалла. По своим морфогенетическим эффектам такие семена напоминают катализаторы химических реакций. Все виды химического морфогенеза, рассмотренные до сих пор, несомненно, агрегативны. В неживых системах явления трансформации гораздо менее распространены. Например, большинство кристаллов не способны превращаться в другие кристаллические формы; они могут расплавиться или раствориться, и тогда их составляющие могут принять участие в других процессах кристаллизации; но это есть разрушение (дезагрегация), за которым следуют другие виды агрегации. Подобным же образом химические реакции включают дезагрегативные и агрегативные изменения. Однако существуют важные примеры молекулярной трансформации, такие как складывание белков, и обратимые изменения формы, которые происходят, когда некоторые ферменты связываются с молекулами, реакции которых они катализируют.[106] Тот факт, что белки складываются гораздо быстрее, чем можно было бы ожидать, если бы они «находили» свою конечную форму путем «случайного поиска», указывает на то, что их складывание происходит по определенным направлениям или по ограниченному числу направлений (раздел 3.3). Эти «канализированные пути изменения» можно рассматривать как хреоды. Чтобы процесс складывания мог начаться, согласно представлениям, излагавшимся выше в разделе 4.1, должен присутствовать морфогенетический зародыш и этот зародыш должен уже иметь характерную трехмерную структуру, которую он имеет в конечной форме белка. Предположение о существовании таких морфогенетических пусковых точек фактически уже было высказано в литературе по складыванию белков: «Высокая скорость раскручивания предполагает необходимость того, чтобы процесс протекал по ограниченному числу направлений, даже когда статистика эта строго ограничена своего рода основными правилами стереохимии, которые содержатся в неявном виде в так называемом графике Рамачандрана. Становится необходимым постулировать в процессе складывания существование ограниченного числа допустимых инициирующих событий. Такие события, обычно называемые нуклеациями, вероятнее всего, происходят в тех частях полипептидной цепи, которые могут участвовать в конформационном равновесии между случайными и кооперативно стабилизированными структурами. Далее важно подчеркнуть, что аминокислотные последовательности полипептидных цепей, предназначенные для того, чтобы служить фабрикой белковых молекул, функционально осмыслены лишь тогда, когда они имеют трехмерную структуру, которая характерна для них в молекуле природного белка. Кажется разумным предположить, что части белковой цепи, которые могут служить как центры нуклеации для складывания, будут теми, которые могут «отклоняться» внутрь и наружу от конформации, которую они имеют в конечном белке, и что они образуют относительно жесткую структуру, стабилизированную набором кооперативных взаимодействий».[107] Такие «центры нуклеации» будут действовать как морфогенетические зародыши через их связывание с морфогенетическим полем белка, которое затем будет канализировать процесс складывания по направлению к характерной конечной форме. 4.3. Морфогенетические поля как вероятностные структуры Орбитали электронов вокруг атомного ядра можно рассматривать как структуры в морфогенетическом поле атома. Эти орбитали могут быть описаны решениями уравнения Шредингера. Однако, согласно квантовой механике, могут быть определены не точные орбиты электронов, но лишь вероятности нахождения электронов в определенных точках; орбитали рассматриваются как распределения вероятностей в пространстве. В контексте гипотезы формативной причинности этот результат предполагает, что так же как эти структуры в морфогенетических полях атомов можно представить как пространственные распределения вероятностей, так и морфогенетические поля вообще не могут быть точно определены, но даются распределениями вероятностей.[108] Мы будем предполагать, что это действительно так, и впредь структуры морфогенетических полей будут считаться вероятностными структурами.[109] Объяснение вероятностной природы этих полей дано в разделе 5.4. Воздействие морфогенетического поля морфической единицы на морфогенетические поля ее частей, которые являются морфическими единицами на низших уровнях (раздел 3.4), можно представить как влияние этой вероятностной структуры высшего уровня на вероятностные структуры низших уровней; поле высшего уровня изменяет (модифицирует) вероятностные структуры полей низших уровней. Следовательно, в процессе морфогенеза поле высшего уровня изменяет вероятность вероятностных событий в морфических единицах низшего уровня, находящихся под его влиянием.[110] В случае свободных атомов электронные события происходят с вероятностями, задаваемыми немодифицированными вероятностными структурами атомных морфогенетических полей. Но когда атомы оказываются в сфере влияния морфогенетического поля молекулы, то есть поля высшего уровня, эти вероятности модифицируются таким образом, что вероятность событий, ведущих к актуализации конечной формы, увеличивается, а вероятность других событий уменьшается. Таким образом, морфогенетические поля молекул ограничивают возможное число атомных конфигураций, ожидаемых на основе вычислений, которые начинаются от вероятностных структур свободных атомов. Именно это и обнаруживается в действительности: например, при складывании белка скорость процесса указывает на то, что система не «исследует» бесчисленные мыслимые конфигурации, в которые могли бы быть организованы атомы (раздел 3.3). Подобным же образом морфогенетические поля кристаллов ограничивают большое число возможных конфигураций, которые могли бы быть разрешены вероятностными структурами составляющих их молекул; в результате при кристаллизации создается одна определенная организация молекул из большого числа других мыслимых структур. Так, морфогенетические поля кристаллов и молекул являются вероятностными структурами в том же смысле, в каком ими являются электронные орбитали в морфогенетических полях атомов. Этот вывод согласуется с известным допущением, что нет качественной разницы между квантово-механическим описанием простых атомных систем и потенциальным квантово-механическим описанием более сложных форм. Но, в отличие от гипотезы формативной причинности, известная теория стремится объяснить сложные системы снизу вверх, то есть через квантово-механические свойства атомов. Различие между этими двумя подходами можно яснее увидеть в историческом контексте. Сама квантовая теория была изначально разработана в связи со свойствами простых систем, таких как атомы водорода. Со временем были введены новые фундаментальные принципы, чтобы объяснить эмпирические наблюдения, например тонкую структуру спектров излучения атомов. Первоначальные квантовые числа, характеризующие дискретные электронные орбитали, были дополнены другим набором чисел, определяющих угловой момент и затем «спин». Последний считается неотъемлемым (нередуцируемым) свойством частиц, таким же как электрический заряд, и имеет собственный закон сохранения. В физике ядерных частиц еще более нередуцируемые факторы, такие как «странность» и «очарование», вместе с дополнительными законами сохранения были введены более или менее специальным образом, чтобы объяснить наблюдения, не объяснимые с помощью уже существовавших квантовых факторов. Более того, открытие большого числа новых субатомных частиц привело к необходимости постулировать все возрастающее число новых видов материальных полей. Когда уже так много новых физических принципов и физических полей было введено для объяснения свойств атомов и субатомных частиц, то распространенное допущение, что на уровнях организации выше атома новые физические принципы или поля не играют роли, кажется весьма произвольным. На самом деле оно есть немногим более нежели реликт атомизма девятнадцатого века; теперь, когда атомы уже более не считаются конечными и неделимыми, его изначальное теоретическое оправдание перестало существовать. С точки зрения гипотезы формативной причинности, хотя существующая квантовая теория, разработанная для описания свойств атомов и субатомных частиц, проливает много света на природу этих морфогенетических полей, она не может быть экстраполирована для описания морфогенетических полей более сложных систем. Нет причин считать, что морфогенетические поля атомов занимают привилегированное положение в системе природы; они являются просто полями морфических единиц на одном определенном уровне сложности. 4.4. Вероятностные процессы в биологическом морфогенезе Есть множество примеров физических процессов, имеющих вероятностные пространственные результаты. В общем случае изменения, включающие нарушения симметрии или гомогенности, являются неопределенными; примеры такого рода можно найти в фазовых переходах между газообразным и жидким, а также жидким и твердым состояниями. Если, например, сферический баллон, наполненный паром, охлажден до температуры ниже точки насыщения в отсутствие внешних градиентов температуры и силы тяжести, жидкость начнет конденсироваться на стенках, но конечное ее распределение будет непредсказуемо и почти никогда не будет сферически симметрично.[111] Термодинамика может предсказать относительные количества жидкости и пара, но не их пространственное распределение. При кристаллизации вещества в однородных условиях пространственное распределение, а также число и размеры кристаллов не могут быть предсказаны; другими словами, если бы тот же самый процесс повторялся в аналогичных условиях, каждый раз пространственный результат различался бы в деталях. Сами формы кристаллов, хотя они демонстрируют определенную симметрию, могут быть неопределенными; хорошо знакомым примером являются снежинки, которые образуют мириады различных форм.[112] В «диссипативных структурах» макроскопических физических и химических систем вдали от термодинамического равновесия случайные флуктуации могут вызвать пространственные картины, например конвекционные ячейки в нагретой жидкости или окрашенные полосы в растворах, где протекает реакция Жаботинского. Математическое описание таких случаев «порядка через флуктуации» методами неравновесной термодинамики дает поразительные аналогии с фазовыми переходами.[113] Эти примеры пространственной неопределенности (индетерминизма) заимствованы из самых простых физических и химических процессов. В живых клетках физико-химические системы гораздо сложнее, чем любые системы в неорганической сфере, и могут включать многие потенциально неопределенные фазовые переходы и неравновесные термодинамические процессы. В протоплазме кристаллическая, жидкая и липидная фазы существуют в динамической взаимосвязи; далее, здесь присутствует также множество видов макромолекул, которые могут объединяться в кристаллические или квазикристаллические агрегаты; липидные мембраны, которые, как «жидкие кристаллы», существуют на границе между жидким и твердым состояниями, подобно коллоидным золям и гелям; электрические потенциалы на мембранах, которые флуктуируют непредсказуемым образом; и «компартменты», содержащие разные концентрации неорганических ионов и других веществ, разделенных мембранами, через которые эти вещества перемещаются вероятностным образом.[114] При такой сложности число энергетически возможных способов пространственного изменения должно быть огромным, и поэтому здесь велик диапазон действия морфогенетических полей путем наложения пространственных ограничений на эти вероятностные процессы. Это не значит, что все формы в живых организмах определяются формативной причинностью. Некоторые конфигурации могут возникать в результате случайных процессов. Другие могут быть полностью объяснены с помощью конфигураций с минимальной энергией: так, сферическую форму свободно плавающих яиц (например, у морских ежей) можно полностью объяснить через поверхностное натяжение клеточной мембраны. Однако весьма ограниченные успехи простых физических объяснений биологических форм[115] предполагают, что большая часть аспектов биологического морфогенеза определяется морфогенетическими полями. Следует вновь подчеркнуть, что эти поля действуют не одни, но вместе с энергетическими и химическими причинами, которые изучают биофизики и биохимики. Пример того, каким способом морфогенетические поля могут оперировать в клетке, дается расположением микротрубочек, тонких палочкообразных структур, образуемых при спонтанной агрегации белковых субъединиц. Микротрубочки играют важную роль микроскопических «подпорок» как в растительных, так и в животных клетках: они контролируют и направляют процессы, такие как клеточное деление (волокна веретена при митозе и мейозе[116] состоят из микротрубочек) и организованное отложение материала клеточной стенки в дифференцирующихся растительных клетках; они служат также внутриклеточными «скелетами», поддерживающими определенные формы клеток, как в радиоляриях.[117] Теперь если пространственное распределение микротрубочек ответственно за организацию многих различных видов процессов и структур внутри клеток, тогда что определяет пространственное распределение самих микротрубочек? Если ответственны другие виды пространственной организации,[118] проблема просто отодвигается на одну ступень назад; а что же определяет сами эти виды организации? Но проблема не может отодвигаться до бесконечности, поскольку развитие эпигенетично, то есть оно включает увеличение пространственного разнообразия и организации, которое не может быть объяснено с помощью предшествующих моделей или структур; рано или поздно что-то другое должно объяснить появление структуры, образующейся при агрегации микротрубочек. Согласно настоящей гипотезе, эта структура объясняется действием специфических морфогенетических полей. Эти поля сильно увеличивают вероятность агрегации микротрубочек в их подходящих положениях либо непосредственно, либо путем установления предшествующего образца соответствующей структуры. Очевидно, структурообразующая активность этих полей зависит от присутствия пересыщенного раствора субъединиц микротрубочек в клетке и от подходящих физико-химических условий для их агрегации; это необходимые условия для формирования микротрубочек, но их самих по себе еще недостаточно для объяснения структуры, которую микротрубочки при этом образуют. Можно возразить, что предполагаемое действие формативной причинности по организации вероятностных процессов в клетках невозможно, поскольку оно привело бы к локальному нарушению второго закона термодинамики. Но это возражение несправедливо. Второй закон термодинамики применим только к ансамблям очень большого числа частиц и к процессам в макроскопическом масштабе. Более того, он приложим лишь к закрытым системам; участок клетки не является закрытой системой, а тем более к таким системам нельзя отнести живые организмы вообще. В живых организмах, как и в области химии, морфогенетические поля иерархически организованы: поля органелл — например, клеточного ядра, митохондрий и хлоропластов — действуют путем упорядочивания в них физико-химических процессов; эти поля подвержены действию полей клеток, которые являются полями более высокого уровня; поля клеток находятся под влиянием полей тканей; поля тканей подчиняются полям органов; а поля органов — полю всего организма как целого. На каждом уровне поля работают путем упорядочивания процессов, которые в противном случае имели бы неопределенный характер. Например, на клеточном уровне морфогенетическое поле организует кристаллизацию микротрубочек и другие процессы, необходимые для координации клеточного деления. Но плоскости, в которых делятся клетки, могут быть неопределенными в отсутствие поля высшего уровня: например, в мозолях растений клетки делятся в более или менее случайных направлениях, создавая хаотическую массу.[119] С другой стороны, в организованной ткани одной из функций ее морфогенетического поля может быть наложение определенной модели организации на плоскости клеточного деления и осуществление таким образом контроля над ростом ткани как целого. Тогда развитие тканей самих по себе может быть неопределенным во многих отношениях, что выявляется, когда они изолируются искусственным путем и выращиваются в культуре;[120] при нормальных условиях эта неопределенность ограничивается полем более высокого уровня — полем органа. Действительно, на каждом уровне в биологических системах, как и в химических, изолированные морфические единицы ведут себя более неопределенно, чем тогда, когда они являются частью морфической единицы более высокого уровня. Морфогенетическое поле ограничивает и организует присущий им индетерминизм. 4.5. Морфогенетические зародыши в биологических системах На клеточном уровне зародыши морфогенетических трансформаций могут быть морфическими единицами низшего уровня в клетках, которые присутствуют как в начале, так и в конце процесса клеточной дифференциации. Возможные морфогенетические зародыши этих трансформаций сразу не очевидны: это могут быть органеллы, макромолекулярные агрегаты, цитоплазматические или мембранные структуры или клеточные ядра. Во многих случаях ядра могут играть эту роль. Но поскольку в одном и том же организме может производиться так много различных типов дифференцированных клеток, если ядра должны действовать как морфогенетические зародыши, они должны быть способны создавать различные модели организации в клетках различных типов: дифференциации клетки должна предшествовать дифференциация ее ядра, благодаря изменениям в его мембране, или в расположении хромосом, или в связях между белками и нуклеиновыми кислотами в хромосомах, или в нуклеолях, или в других компонентах. Такие изменения могут быть вызваны непосредственно или опосредованно, через воздействие морфогенетического поля высшего уровня — поля дифференцирующейся ткани. Действительно, имеется немало данных о том, что многим типам клеточной дифференциации предшествуют изменения ядер. Выдвигаемое здесь предположение отличается от обычной интерпретации таких изменений тем, что в нем эти изменения рассматриваются не как просто химические, обусловленные образованием специальных видов РНК-мессенджера (предвестник), но, в дополнение к этому, как морфогенетические: модифицированные ядра могут служить как зародыши, с которыми ассоциируются (связываются) специфические морфогенетические поля дифференцированных клеток.[121] Существует по крайней мере один процесс клеточного морфогенеза, в котором ядро не может служить морфогенетическим зародышем: это деление ядер. Ядро теряет свою идентичность как отдельная структура, когда ядерная мембрана разрушается и исчезает.[122] Двойные, сильно скрученные хромосомы выстраиваются в линии в экваториальной области митотического веретена,[123] и затем полный набор хромосом движется к каждому из полюсов веретена. Затем вокруг каждого набора хромосом нарастает новая ядерная мембрана, в результате чего образуются дочерние ядра. Морфогенетическими зародышами для этих процессов должны быть экстра-нуклеарные (внеядерные) структуры или органеллы, и таких зародышей должно быть два.[124] Развитие тканей и органов обычно включает как трансформативные, так и агрегативные изменения. В этих морфогенезах морфогенетическими зародышами должны быть клетки или группы клеток, которые присутствуют и как часть конечной формы, и в начале морфогенетического процесса; это не могут быть те специализированные клетки, которые появляются только после того, как процесс начался. Таким образом, здесь морфогенетическими зародышами, вероятно, могут быть только относительно слабо специализированные клетки, которые изменяются мало. В высших растениях такие клетки присутствуют, например в апикальных зонах меристем[125] или точек роста.[126] У побегов стимул к цветению трансформирует меристемы таким образом, что они порождают, скорее, цветы, нежели листья и другие вегетативные структуры; апикальные зоны, должным образом модифицированные стимулом к цветению, могут быть морфогенетическими зародышами для такой трансформации. В эмбрионах животных эмбриологами уже было обнаружено множество «организующих центров», которые играют ключевую роль в развитии тканей и органов; одним из примеров является апикальный гребень эктодермы[127] на верхушках развивающихся зачатков конечностей.[128] Эти «организующие центры» вполне могут быть зародышами, с которыми связываются морфогенетические поля более высоких уровней. Хотя как в химии, так и в биологии для описания явлений можно предложить морфогенетические термины, а иногда и идентифицировать соответствующие структуры, все же многое остается неясным, особенно происхождение индивидуальной формы каждого морфогенетического поля, а также способа его соединения со своим зародышем. Рассмотрение этих проблем в следующей главе приводит нас к более сложной гипотезе формативной причинности, которую, хотя она и выглядит удивительно и непривычно, понять оказывается легче. Примечания:1 Особенно четкое представление дано у Monod (1972). 10 Например, Elsasser (1966, 1975); von Bertalanfi (1971). 11 См., например, дискуссию между Уоддингтоном и Р. Томом в Waddington (ed.) (1969), p. 42. 12 Это обсуждается в последней главе настоящей книги. 105 Отождествление морфогенетических полей с электромагнитными полями привело к большой путанице в теории электродинамических «полей жизни» X. С. Бэра (Burr, 1972). Он приводит неопровержимые свидетельства того, что живые организмы связаны с электромагнитными полями, которые изменяются с изменением организмов, но затем начинает утверждать, что эти поля управляют морфогенезом, действуя как «проекты» развития, что уже совсем другое дело. 106 Обзор литературы по конформационным изменениям белков в растворе дан у Вильямса (Williams, 1979). 107 Anfinsen (1973), p. 228. 108 Общее обсуждение вероятностной причинности дано в Supps (1970). 109 Сравните с концепцией полей вероятности, или склонности (propensity), сэра Карла Поппера (Popper, 1967; Popper and Eccles, 1977). 110 Это предположение согласуется с подходом в квантовой физике, развиваемым Бомом и Хайли (Bohm, 1969, 1980 and Hiley, 1980). 111 Этот и другие примеры того, что Р. Том называет «обобщенными катастрофами», обсуждаются в главе 6 его книги «Структурная стабильность и морфогенез». 112 Benley and Humphreys (1962). 113 Nicolis и Prigogine (1977). Другой, но родственный подход к этим проблемам описан у Хакена (Haken, 1977). 114 Stevens (1977). 115 Сэр Д'Арси Томпсон в своем классическом эссе «О росте и форме» («On Growth and Form», 1942) предположил, что многие аспекты биологического морфогенеза могут быть объяснены на языке физических сил: например, план клеточного деления — на языке поверхностного натяжения, которое будет стремиться достичь минимальной площади поверхности. Но существует так много исключений, что эти простые интерпретации имели небольшой успех. Обсуждение теорий Томпсона можно найти у Медавара (Medavar, 1968). 116 Митоз (от греч. нить) — непрямое деление ядра клетки и ее тела; в ходе такого деления возникают последовательные характерные фазы с чет кой морфологической картиной. Мейоз (от греч. уменьшение, убывание) — процесс деления созревающих половых клеток. — Прим. ред. 117 Объяснение свойств и функций микротрубочек было недавно дано в Dustin (1978) и Roberts and Hyams (eds.) (1979). 118 Одно предположение состоит в том, что глад кий эндоплазматический ретикулум играет роль в транспорте субъединиц микротрубочек к участкам, где они агрегируют (Burgess и Northcote, 1968). Предполагалось также сочетание «образующих ядра элементов», которые могут быть или не быть связаны воедино в «организующих центрах микротрубочек» (J. В. Tucker in Roberts and Hyams (eds.), 1979). 119 Streern Henshaw(1965). 120 Примеры даны у Вилмера (Willmer, 1970). 121 В некоторых случаях ядра разрушаются на последних стадиях дифференциации (например, ксилемные сосуды у растений, красные кровяные тельца у млекопитающих). В этих случаях ядра могут действовать как морфогенетические зародыши для процесса дифференциации до того момента, пока они остаются нецикличными; затем конечные стадии дифференциации протекают чисто механически с помощью прямых химических процессов, не подверженных морфогенетическому упорядочиванию, через выброс гидролитических ферментов. 122 У некоторых водорослей, например Oedogonium, ядерная мембрана в течение митоза остается нецикличной; возможно, это особенность примитивной эволюции (Pickett-Heaps, 1975). 123 Митотическое веретено (веретено деления) — образуется в делящейся клетке в период определенных фаз (после исчезновения ядерной оболочки) и занимает до половины объема клетки; временное тело нуклепротеидной при роды; обеспечивает расхождение хромосом в митозе и мейозе. — Прим. ред. 124 У животных центриоли могут оказаться вероятными кандидатами на эту роль, но у высших растений нет центриолей. В обоих случаях «организующие центры микротрубочек» могут быть ответственны за развитие полюсов веретена; центриоли могут быть просто «пассажирами», для которых гарантировано одинаковое попадание в дочерние клетки путем связывания с этими центрами (Pickett-Heaps, 1969). Центриоли служат организующими центрами, или морфогенетическими зародышами, для развития ресничек и бичей (flagella). 125 Меристема (от греч. делимый), или точка рос та, — группа клеток, способных к активному клеточному делению. Такие клетки имеются, напри мер, в конце корня растения под корневым чех- ликом, который защищает их от повреждений и облегчает продвижение корня в почве во время роста. — Прим. пер. 126 Clowes (1961). 127 Эктодерма — 1) наружный зародышевый лис ток эмбриона многоклеточных животных; 2) наружный слой двуслойных животных — губок, кишечнополостных. — Прим. ред. 128 Wolpert (1978). |
|
||
|
Главная | Контакты | Нашёл ошибку | Прислать материал | Добавить в избранное |
||||
|
|
||||